Thực vật là những sinh vật có khả năng tạo cho mình chất dinh dưỡng từ những hợp chất vô cơ đơn giản và xây dựng thành những phần tử phức tạp nhờ quá trình quang hợp, diễn ra trong lục lạp của thực vật. Như vậy thực vật chủ yếu là các sinh vật tự dưỡng. Quá trình quang hợp sử dụng năng lượng ánh sáng được hấp thu nhờ sắc tố màu lục - Diệp lục có ở tất cả các loài thực vật (không có ở động vật) và nấm là một ngoại lệ, dù không có chất diệp lục nhưng nó thu được các chất dinh dưỡng nhờ các chất hữu cơ lấy từ sinh vật khác hoặc mô chết. Thực vật còn có đặc trưng bởi có thành tế bào bằng xenluloza (không có ở động vật). Thực vật không có khả năng chuyển động tự do ngoại trừ một số thực vật hiển vi có khả năng chuyển động được. Thực vật còn khác ở động vật là chúng phản ứng rất chậm với sự kích thích, sự phản ứng lại thường phải đến hàng ngày và chỉ trong trường hợp có nguồn kích thích kéo dài.
Thực vật là một nhóm chính các sinh vật, bao gồm các sinh vật rất quen thuộc như cây gỗ, cây hoa, cây cỏ, dương xỉ hay rêu. Khoảng 350.000 loài thực vật, được xác định như là thực vật có hạt, rêu, dương xỉ và các dạng gần giống như dương xỉ, đã được ước tính là đang tồn tại. Vào thời điểm năm 2004, khoảng 287.655 loài đã được nhận dạng, trong đó 258.650 loài là thực vật có hoa và 15.000 loài rêu.
Aristotle phân chia sinh vật ra thành thực vật, nói chung là không di chuyển được, và động vật. Trong hệ thống của Linnaeus, chúng trở thành các giới Vegetabilia (sau này là Plantae) và Animalia. Kể từ đó trở đi, một điều trở nên rõ ràng là giới thực vật như trong định nghĩa nguyên thủy đã bao gồm vài nhóm không có quan hệ họ hàng gì, và người ta đã loại nấm và một vài nhóm tảo ra để tạo thành các giới mới. Tuy nhiên, chúng vẫn còn được coi là thực vật trong nhiều ngữ cảnh. Thực vậy, bất kỳ cố gắng nào nhằm làm cho "thực vật" trở thành một đơn vị phân loại đơn duy nhất đều chịu một số phận bi đát, do thực vật là một khái niệm được định nghĩa một cách gần đúng, không liên quan với các khái niệm được cho là đúng của phát sinh loài, mà phân loại học hiện đại đang dựa vào nó.
Sự đa dạng
Hơn 350.000 loài thực vật, gồm thực vật có hạt, rêu, dương xỉ và cận dương xỉ (fern ally) được thống kê hiện đang tồn tại. Năm 2004, 287.655 loài được xác định, trong số đó 258.650 là loài có hoa, 16.000 loài rêu, 11.000 loài dương xỉ và 8.000 loài tảo xanh.
Thực vật có phôi
Quen thuộc nhất là các loài thực vật đa bào sống trên mặt đất, được gọi là thực vật có phôi (Embryophyta). Chúng bao gồm các loài thực vật có mạch, là các loại thực vật với các hệ thống đầy đủ của lá, thân và rễ. Chúng cũng bao gồm cả một ít các loài có quan hệ họ hàng gần với thực vật có mạch, thường được gọi trong khoa học là Bryophyta, với các loài rêu là phổ biến nhất.
Tất cả các loại thực vật này đều có các tế bào nhân chuẩn với các màng tế bào được tạo thành từ xenluloza và phần lớn thực vật thu được nguồn năng lượng thông qua quang hợp, trong đó chúng sử dụng ánh sáng và điôxít cacbon để tổng hợp thức ăn. Khoảng 300 loài thực vật không quang hợp mà sống ký sinh trên các loài thực vật quang hợp khác. Thực vật là khác với tảo lục, mà chúng đã tiến hóa từ đó, ở điểm là chúng có các cơ quan sinh sản chuyên biệt được các mô không sinh sản bảo vệ.
Các loài rêu trong nhóm Bryophyta lần đầu tiên xuất hiện từ đầu đại Cổ Sinh. Chúng chỉ có thể sống sót trong các môi trường ẩm ướt, và giữ nguyên kích thước nhỏ trong suốt chu trình sống của chúng. Nó bao gồm sự luân phiên giữa hai thế hệ: giai đoạn đơn bội, được gọi là thể giao tử và giai đoạn lưỡng bội, được gọi là thể bào tử. Thể bào tử có thời gian sống ngắn và là phụ thuộc vào cha, mẹ của chúng.
Thực vật có mạch xuất hiện lần đầu tiên vào thời kỳ của kỷ Silur (409-439 Ma), và vào kỷ Devon (359-416 Ma) chúng đã đa dạng hóa và lan rộng trong nhiều môi trường đất khác nhau. Chúng có nhiều cơ chế thích nghi, cho phép chúng vượt qua các hạn chế của Bryophyta. Các cơ chế này bao gồm lớp biểu bì (chất cutin) chống bị khô và các mô có mạch để vận chuyển nước trong khắp cơ thể. Ở nhiều loài, thể bào tử đóng vai trò như một cá thể tách rời, trong khi thể giao tử vẫn là nhỏ.
Thực vật có hạt nguyên thủy đầu tiên, Pteridospermatophyta (dương xỉ có hạt) và nhóm Cordaitales, cả hai nhóm này hiện nay đã tuyệt chủng, đã xuất hiện vào cuối kỷ Devon và đa dạng hóa trong kỷ Than Đá (280-340 Ma), với sự tiến hóa kế tiếp diễn ra trong kỷ Permi (248-280 Ma) và kỷ Trias (200-251 Ma). Ở chúng, giai đoạn thể giao tử bị suy giảm hoàn toàn, và thể bào tử bắt đầu cuộc sống bên trong lớp bao bọc, gọi là hạt, chúng phát triển khi đang ở trên thực vật cha mẹ và với sự thụ phấn bằng các hạt phấn. Trong khi các loài thực vật có mạch khác, chẳng hạn như dương xỉ, sinh sản nhờ các bào tử và cần có sự ẩm ướt để phát triển thì một số thực vật có hạt có thể sinh sống và sinh sản trong các điều kiện cực kỳ khô cằn.
Các loài thực vật có hạt đầu tiên được nói đến như là thực vật hạt trần (Gymnospermae), do phôi hạt không được bao bọc trong một cấu trúc bảo vệ khi thụ phấn, với các hạt phấn trực tiếp hạ xuống phôi. Bốn nhóm còn sống sót hiện vẫn phổ biến rộng khắp, cụ thể là thực vật quả nón, là nhóm cây thân gỗ thống trị trong một vài quần xã sinh vật. Thực vật hạt kín (Angiosperm), bao gồm thực vật có hoa, là nhóm thực vật chính cuối cùng đã xuất hiện, nảy ra từ thực vật hạt trần trong kỷ Jura (146-200 Ma) và đa dạng hóa nhanh chóng trong kỷ Phấn Trắng (65-146 Ma). Chúng khác với thực vật hạt trần ở chỗ các phôi hạt được bao bọc, vì thế phấn hoa cần phải phát triển một ống để xâm nhập qua lớp vỏ bảo vệ hạt; chúng là nhóm thống trị trong giới thực vật ngày nay ở phần lớn các quần xã sinh vật.
Phát sinh loài
Phát sinh loài dưới đây của Plantae lấy theo Kenrick và Crane[10], với biến đổi đối với Pteridophyta lấy theo Smith và ctv.[11]. Prasinophyceae có thể là nhóm cơ sở cận ngành đối với toàn bộ thực vật xanh.
Tảo
Tảo bao gồm vài nhóm sinh vật khác biệt, sinh ra nguồn năng lượng thông qua quang hợp. Dễ thấy nhất là các loài tảo biển, là các loại tảo đa bào thông thường rất giống với thực vật trên đất liền, được tìm thấy bao gồm tảo lục, tảo đỏ và tảo nâu. Các nhóm tảo này cùng với các nhóm tảo khác cũng bao gồm các sinh vật đơn bào khác nhau.
Thực vật có phôi đã phát triển và tiến hóa từ tảo lục; cả hai được gọi tổng thể như là thực vật xanh (Viridaeplantae). Giới thực vật (Plantae) hiện nay thông thường được chọn lựa sao cho nó là một nhóm đơn ngành, như chỉ ra trên đây. Với một ít ngoại lệ trong nhóm tảo lục, tất cả các dạng này đều có màng tế bào chứa xenluloza và lạp lục chứa các chất diệp lục a và b, và lưu trữ nguồn thức ăn dưới dạng tinh bột. Chúng trải qua sự phân bào có tơ khép kín mà không có các trung thể, và thông thường có các ti thể với các nếp màng trong thể sợi hạt phẳng.
Các lạp lục của thực vật xanh cũng được 2 màng bao quanh, gợi ý rằng chúng có nguồn gốc trực tiếp từ vi khuẩn lam nội cộng sinh. Điều này cũng đúng với tảo đỏ (xem Archaeplastida), và hai nhóm này nói chung được coi là có nguồn gốc chung. Ngược lại, phần lớn các nhóm tảo khác có các lạp lục với 3 hoặc 4 màng. Về tổng thể chúng là không có quan hệ họ hàng gần gũi với thực vật xanh, có lẽ có được các lạp lục tách rời khỏi các nhóm tảo lục hay tảo đỏ cộng sinh.
Không giống như thực vật có phôi và tảo, nấm không có cơ chế quang hợp, mà là dạng sinh vật hoại sinh: chúng thu được nguồn thức ăn bằng cách phân hủy và hấp thụ các vật chất xung quanh chúng. Phần lớn các loài nấm được tạo thành bởi các ống cực nhỏ, gọi là sợi nấm, chúng có thể hoặc không thể phân chia thành các tế bào nhưng chứa nhân tế bào. Phần giống như quả, trong đó các loài nấm đất là thông thường nhất, trên thực tế chỉ là các cấu trúc sinh sản của nấm. Chúng không có quan hệ tới bất kỳ nhóm thực vật quang hợp nào, mà có quan hệ họ hàng gần gũi với động vật.
Tầm quan trọng
Quang hợp và cố định điôxít cacbon của thực vật có phôi và tảo là nguồn năng lượng cũng như nguồn các chất hữu cơ cơ bản nhất trong gần như mọi môi trường sống trên Trái Đất. Quá trình này cũng làm thay đổi hoàn toàn thành phần của khí quyển Trái Đất, với kết quả là nó có thành phần ôxy cao. Động vật và phần lớn các sinh vật khác là các sinh vật hiếu khí, phụ thuộc vào ôxy; chúng không thể sinh sống được trong các môi trường hiếm khí.
Phần lớn nguồn dinh dưỡng của loài người phụ thuộc vào ngũ cốc. Các loại thực vật khác mà con người cũng dùng bao gồm các loại hoa quả, rau, gia vị và cây thuốc. Một số loài thực vật có mạch, được coi là cây thân gỗ hay cây bụi, sản sinh ra các thân gỗ và là nguồn vật liệu xây dựng quan trọng. Một số các loài cây khác được sử dụng với mục đích làm cảnh hay trang trí, bao gồm nhiều loại cây hoa.
Như vậy, có thể cho rằng thực vật là yếu tố cơ bản của sự sống trên Trái Đất. Không có thực vật thì nhiều sinh vật khác cũng không thể tồn tại, vì các dạng sinh vật cao hơn đều trực tiếp hoặc gián tiếp phụ thuộc vào thực vật và về cơ bản đều sử dụng thực vật như là nguồn thức ăn. Trong khi đó, hầu hết mọi thực vật đều có thể sử dụng ánh sáng Mặt Trời tự tạo thức ăn cho mình.
Quan hệ sinh thái
Cây nắp ấm Nepenthes villosa, một loài thực vật ăn thịt.
Quang hợp do thực vật đất liền và rong, tảo thực hiện là nguồn năng lượng và vật chất hữu cơ cơ bản trong gần như mọi hệ sinh thái. Quá trình quang hợp đã làm thay đổi căn bản thành phần của khí quyển Trái Đất thời nguyên thủy, với kết quả là 21% ôxy như ngày nay. Động vật và phần lớn các sinh vật khác là các sinh vật hiếu khí, phụ thuộc vào ôxy; những sinh vật không hiếu khí là những loài có môi trường sống bị giam hãm trong các môi trường kỵ khí. Thực vật là các nhà sản xuất chính trong phần lớn các hệ sinh thái mặt đất và tạo thành nền tảng của chuỗi thức ăn trong các hệ sinh thái này. Nhiều động vật dựa vào thực vật như là nơi trú ẩn cũng như nguồn thức ăn và ôxy.
Thực vật đất liền là thành phần quan trọng trong chu trình nước và một vài chu trình hóa địa sinh khác. Một số thực vật cộng sinh cùng với các vi khuẩn cố định đạm, làm cho thực vật trở thành một phần quan trọng trong chu trình nitơ. Các rễ thực vật đóng vai trò thiết yếu trong sự hình thành và phát triển của các loại đất và ngăn cản xói mòn đất. Các quần xã sinh vật trên Trái Đất được gọi tên theo loại thực vật là do thực vật là các sinh vật thống lĩnh trong các quần xã này.
Hàng loạt các động vật đã cùng tiến hóa với thực vật. Nhiều động vật thụ phấn cho hoa để đổi lấy là nguồn thức ăn trong dạng phấn hoa hay mật hoa. Nhiều động vật cũng làm các hạt được phân tán rộng khắp do chúng ăn quả và để lại hạt trong phân của chúng. Cây ổ kiến gai (Myrmecodia armata) là những thực vật đã cùng tiến hoa với kiến. Cây cung cấp nơi cư trú, và đôi khi là thức ăn cho kiến. Để đổi lại, kiến bảo vệ cây tránh khỏi các loài động vật ăn cỏ và đôi khi là các loài cây cạnh tranh khác. Các chất thải của kiến lại cung cấp một lượng phân bón hữu cơ cho cây.
Phần lớn các loài thực vật gắn liền với nhiều loại nấm tại hệ rễ của chúng, trong dạng cộng sinh phụ thuộc, được biết đến như là nấm rễ (mycorrhiza). Nấm giúp cho cây thu được nước và các chất dinh dưỡng từ đất, trong khi cây cung cấp cho nấm các loại cacbohyđrat được sản xuất nhờ quang hợp. Một số thực vật còn là nơi ở cho các loại nấm sống trên cây, chúng bảo vệ cây khỏi các loài ăn cỏ bằng cách tiết ra các chất có độc tính. Một loại nấm như vậy là Neotyphodium coenophialum, có trên những cây cỏ đuôi trâu cao (Festuca arundinacea) đã gây ra nhiều tổn thất kinh tế cho ngành chăn nuôi bò của Hoa Kỳ.
Các dạng khác nhau của sự sống ký sinh cũng khá phổ biến giữa các loài thực vật, từ dạng bán ký sinh như cây tầm gửi (một phần bộ Santalales) chỉ đơn thuần lấy đi một số chất dinh dưỡng từ cây chủ và vẫn có các lá có khả năng quang hợp, tới các loài ký sinh hoàn toàn như các loài cỏ chổi (chi Orobanche) hay các loài cỏ thuộc chi Lathrea lấy tất cả các loại chất dinh dưỡng chúng cần thông qua sự kết nối vào rễ các loài thực vật khác, và không có diệp lục. Một số loài thực vật, được biết đến như là dị dưỡng nấm, chúng ký sinh các loài nấm rễ, và vì thế có cơ chế hoạt động ký sinh ngoài trên các loài thực vật khác.
Nhiều loài thực vật là biểu sinh, nghĩa là chúng sống trên các loài thực vật khác, thường là trên các cây thân gỗ, mà không ký sinh các cây này. Thực vật biểu sinh có thể gián tiếp gây hại cho cây chủ bằng cách ngăn chặn nguồn chất khoáng và ánh sáng mà nếu không có chúng thì cây chủ đã nhận được. Một lượng lớn thực vật biểu sinh có thể làm gãy các cành cây to. Nhiều loài lan, dứa, dương xỉ và rêu thường có kiểu sống này.
Một số ít loài thực vật lại là cây ăn thịt, chẳng hạn như bẫy ruồi Venus (Dionaea muscipula) và các loài gọng vó. Chúng bẫy các loài động vật nhỏ và phân hủy con mồi để hấp thụ các khoáng chất, đặc biệt là nitơ.
Một số loài thực vật có cơ chế bảo vệ đặc biệt, chẳng hạn các gai trên thân cây mâm xôi.
Những thực vật đơn giản như tảo có thể có khoảng thời gian sống ngắn khi tính theo từng cá thể, nhưng các quần thể tảo nói chung có tính chất theo mùa. Các loại thực vật khác có thể được sắp xếp theo kiểu phát triển mang tính theo mùa của chúng thành:
Cây một năm: Sống và sinh sản trong một mùa sinh trưởng.
Cây hai năm: Sống trong hai mùa sinh trưởng; thường sinh sản vào năm thứ hai.
Cây lâu năm: Sống nhiều mùa sinh trưởng; liên tục sinh sản khi đã trưởng thành.
Trong số các thực vật có mạch, cây lâu năm bao gồm cả cây thường xanh, chúng giữ lá trong cả năm, và cây lá sớm rụng, thường rụng lá trên một số phần nhất định. Ở những vùng có khí hậu ôn đới và phương bắc, nói chung chúng bị rụng lá khi mùa đông tới; nhiều loài thực vật miền nhiệt đới rụng lá vào mùa khô.
Tốc độ tăng trưởng của thực vật nói chung là rất khác nhau. Một số loại rêu lớn chậm hơn 1 μm/h, trong khi phần lớn các cây thân gỗ đạt 25-250 μm/h. Một số loài dây leo, chẳng hạn sắn dây, không cần sản sinh ra các mô hỗ trợ dày, có thể tăng trưởng tới 12.500 μm/h.
Hóa thạch
Các hóa thạch thực vật, bao gồm rễ, gỗ, lá, hạt, quả, phấn hoa, bào tử và hổ phách (nhựa hóa thạch do một số loài thực vật sinh ra). Hóa thạch của thực vật sống trên đất liền được ghi nhận lại trong các trầm tích đất liền, sông, hồ và ven biển. Các phấn hoa, bào tử và tảo (Dinoflagellata và Acritarch) được sử dụng để xác định niên đại các tầng đá trầm tích. Các phần còn lại của thực vật hóa thạch là không phổ biến như của động vật, mặc dù các hóa thạch thực vật là khá phổ biến mang tính cục bộ trong nhiều khu vực trên thế giới.
Các thực vật hóa thạch sớm nhất được biết đến từ kỷ Devon, bao gồm đá phiến silic Rhynie tại Aberdeenshire, Scotland. Các mẫu được bảo quản tốt nhất, mà từ đó kết cấu tế bào của chúng đã được miêu tả, được tìm thấy trong khu vực này. Sự bảo quản hoàn hảo đến mức các phần của các thực vật cổ này chỉ rõ từng tế bào riêng biệt trong mô thực vật. Kỷ Devon cũng cho thấy sự tiến hóa của những thực vật mà nhiều người tin là của loại cây thân gỗ hiện đại đầu tiên, Archaeopteris. Cây này giống như dương xỉ và có thân gỗ và lá lược của dương xỉ, không sinh ra hạt.
Các lá hóa thạch của bạch quả từ kỷ Jura tại Anh.
Các đơn vị than đá là nguồn chính của hóa thạch thực vật thuộc đại Cổ Sinh, với nhiều nhóm thực vật đã tồn tại vào thời kỳ này. Các đống đổ nát trong các mỏ than là các khu vực tốt nhất để thu thập; than tự bản thân nó là các phần còn lại của thực vật hóa thạch, mặc dù các chi tiết cấu trúc của các hóa thạch thực vật là ít rõ ràng trong than. Trong rừng hóa thạch tại công viên Victoria ở Glasgow, Scotland, các gốc cây của nhóm thực vật Lepidodendron được tìm thấy ở các vị trí phát triển nguyên thủy của chúng.
Các phần hóa thạch của thực vật quả nón và thực vật hạt kín như rễ, thân và cành có thể khá phổ biến trong các lớp đá trầm tích trong các hồ và ven bờ từ đại Trung Sinh và đại Tân Sinh. Tùng đỏ duyên hải (chi Sequoia) và các liên minh của nó như mộc lan, sồi và các loài cọ cũng thường được tìm thấy.
Gỗ hóa đá cũng khá phổ biến ở một số nơi trên thế giới, chủ yếu tìm thấy trong các khu vực khô cằn và sa mạc, những nơi chúng hay bị lộ thiên sớm do xói mòn. Gỗ hóa đá thường chứa nhiều silic (các chất hữu cơ bị thay thế bằng điôxít silic), và các mô thụ phấn thường được bảo quản khá chi tiết. Các mẫu vật như thế có thể cắt và đánh bóng bằng các dụng cụ chạm trổ đá. Các rừng hóa thạch chứa gỗ hóa đá đã được tìm thấy ở mọi châu lục.
Các hóa thạch của dương xỉ có hạt như Glossopteris được phân bổ khá rộng rãi ở vài châu lục thuộc Nam bán cầu, một thực tế hỗ trợ ý tưởng ban đầu của Alfred Wegener về thuyết trôi dạt lục địa.
Cơ chế của quá trình quang hợp
Quá trình quang hợp chia làm hai pha: Pha sáng và pha tối:
Pha sáng của quang hợp: Chỉ xảy ra khi có ánh sáng và diễn ra trên các hạt grana. Pha sáng có hai giai đoạn:
Giai đoạn quang lí: Là giai đoạn hấp thu năng lượng ánh sáng nhờ hoạt động của các phân tử sắc tố quang hợp gọi chung là diệp lục và chuyển năng lượng giữa các sắc tố.
Năng lượng ánh sáng hấp thu bởi các sắc tố khác sẽ được chuyển tới diệp lục a và bản thân phân tử diệp lục a hấp thu năng lượng ánh sáng. Sau khi hấp thu năng lượng ánh sáng, phân tử diệp lục ở trạng thái kích động (kí hiệu là DL*), dồi dào năng lượng.
Giai đoạn quang hoá: Là giai đoạn chỉ sử dụng năng lượng photon hấp thu được vào các phản ứng quang hoá để hình thành nên các hợp chất dự trữ năng lượng và các hợp chất khử. Bao gồm quá trình quang hoá khởi nguyên, quá trình quang phân li nước và quá trình photphoril hoá quang hoá. Các quá trình đó được thực hiện cùng với dòng vận chuyển điện tử vòng và không vòng
- Dòng vận chuyển điện tử vòng: Điện tử từ diệp lục qua chuỗi truyền điện tử, sau đó lại quay về diệp lục và trong quá trình truyền điện tử ATP được tổng hợp.
Dòng vận chuyển điện tử không vòng Sản phẩm của pha sáng: ATP, NADPH2, O2b.Pha tối của quá trình quang hợp
Pha tối của quang hợp diễn ra cả khi có ánh sáng và trong tối tại stroma. Pha tối sử dụng sản phẩm của pha sáng (ATP, NADPH2), các enzim trong stroma và đường ribozơ 1,5đi (P) để cố định CO2.
Như vậy, để khử ba phân tử CO2 cần 9ATP và 6NADPH2, tạo ra một phân tử C3 (glixeraldehit chứa liên kết cao năng)
Thực vật có phôi (Embryophyta) là nhóm phổ biến và quen thuộc nhất của thực vật. Chúng bao gồm cây gỗ, cây hoa, dương xỉ, rêu và nhiều loại cây xanh đất liền khác. Tất cả đều là sinh vật nhân chuẩn đa bào phức tạp với các cơ quan sinh sản chuyên biệt. Với một số ít ngoại lệ, thực vật có phôi nói chung thu được năng lượng từ quang hợp (nghĩa là bằng cách hấp thụ ánh sáng); và chúng tổng hợp thức ăn cho mình từ điôxít cacbon. Thực vật có phôi có thể phân biệt với tảo đa bào có sử dụng diệp lục ở chỗ chúng có các mô vô sinh trong các cơ quan sinh sản. Ngoài ra, thực vật có phôi chủ yếu thích nghi với cuộc sống trên đất liền, mặc dù một số loài là thủy sinh thứ cấp. Vì thế, đôi khi người ta còn gọi thực vật có phôi là thực vật đất liền hay thực vật trên đất liền.
Người ta cho rằng thực vật có phôi đã phát triển lên từ các tảo lục phức tạp (Chlorophyta) trong đại Cổ Sinh. Nhóm Charales hay luân tảo dường như là minh họa còn sinh tồn tốt nhất cho bước phát triển này. Các thực vật tương tự như tảo này phải trải qua quá trình luân phiên giữa thể đơn bội và lưỡng bội trong các thế hệ (tương ứng là thể giao tử và thể bào tử). Tuy nhiên, ở các thực vật có phôi đầu tiên thì thể bào tử đã rất khác biệt về cấu trúc và chức năng, mặc dù vẫn duy trì ở dạng nhỏ và phụ thuộc vào cha mẹ trong suốt cuộc đời ngắn của nó. Những thực vật như vậy được gọi là 'thực vật không mạch' hay 'rêu'. Chúng bao gồm các nhóm còn sinh tồn sau:
Bryophyta (rêu thật sự)
Anthocerotophyta (rêu sừng)
Marchantiophyta (rêu tản)
Tất cả các nhóm rêu nói trên là tương đối nhỏ về kích thước và thông thường gắn liền với các môi trường ẩm ướt, dựa vào nước để phát tán các bào tử của chúng.
Các thực vật khác, thích nghi tốt hơn với các điều kiện trên đất liền, đã xuất hiện trong kỷ Silur. Trong kỷ Devon, chúng đã đa dạng hóa và lan tỏa tới nhiều môi trường đất khác nhau, trở thành thực vật có mạch (Tracheophyta). Thực vật có mạch có các mô mạch (quản bào), có chức năng chuyên chở nước trong cơ thể và có lớp biểu bì (lớp cutin) để ngăn chặn sự thoát nước và giữ ẩm. Ở phần lớn thực vật có mạch, thể bào tử là các thành phần thống lĩnh và chúng phát triển thành các lá, thân và rễ, trong khi thể giao tử vẫn là rất nhỏ và ít.
Tuy nhiên, nhiều thực vật có mạch vẫn phát tán bằng bào tử. Chúng bao gồm 2 nhóm còn sinh tồn là:
Lycopodiophyta (thông đất, thạch tùng)
Pteridophyta (dương xỉ, mộc tặc)
Các nhóm khác, lần đầu tiên xuất hiện vào cuối đại Cổ Sinh, sinh sản bằng cách sử dụng các nang có khả năng giữ ẩm, được gọi là hạt. Các nhóm này được gọi là tương ứng là thực vật có hạt (Spermatophyta). Ở các dạng này, thể giao tử bị suy giảm hoàn toàn, tạo ra các dạng đơn tế bào gọi là phấn và trứng, trong khi thể bào tử bắt đầu cuộc sống của nó là nằm trong hạt. Một số thực vật có hạt có thể sống sót trong các điều kiện cực kỳ khô cằn chứ không giống như các tiền bối của nó là dạng gắn liền với nước nhiều hơn. Thực vật có hạt bao gồm các nhóm còn sinh tồn sau:
Cycadophyta (Tuế)
Ginkgophyta (Bạch quả)
Pinophyta (Thông)
Gnetophyta (Dây gắm)
Magnoliophyta (Thực vật có hoa)
Bốn nhóm đầu tiên được gọi chung là thực vật hạt trần, do thể bào tử phôi mầm của chúng không được bao bọc cho đến sau khi thụ phấn. Ngược lại, trong số thực vật có hoa hay thực vật hạt kín thì phấn hoa đã phát triển một ống để thâm nhập vào lớp áo hạt. Thực vật hạt kín là nhóm lớn của thực vật đã xuất hiện sau cùng, phát triển lên từ thực vật hạt trần trong thời gian của kỷ Jura, và sau đó lan truyền nhanh chóng trong kỷ Phấn Trắng. Ngày nay, chúng là nhóm thực vật thống lĩnh nhất trong quần xã sinh vật đất liền.
Lưu ý rằng phân loại thực vật ở cấp bậc cao là biến động một cách đáng kể. Một số tác giả đã hạn chế giới Plantae là chỉ bao gồm Embryophyta, nhưng các tác giả khác thì lại đặt cho chúng nhiều loại tên và cấp bậc. Các nhóm liệt kê ở đây là các ngành thông thường hay được đề cập tới, nhưng cũng có thể chỉ được coi là các lớp, và đôi khi chúng bị dồn lại thành ít ngành hơn (tới 2). Một số phân loại coi thuật ngữ Embryophyta như là nhóm ở cấp siêu ngành, bao gồm các thực vật đất liền và một số loài trong nhóm Charophyceae trong phân giới được đặt tên là Streptophyta.
Ở cấp độ hiển vi, các tế bào của thực vật có phôi vẫn là tương tự như các tế bào ở tảo lục. Chúng là các tế bào nhân chuẩn, với thành tế bào bao gồm xenluloza và các thể hạt bao quanh bởi hai lớp màng. Chúng thông thường có dạng là lạp lục, tiến hành quang hợp và lưu trữ nguồn dự trữ thức ăn dưới dạng tinh bột với đặc trưng là có các sắc tố dưới dạng các diệp lục a và b, nói chung làm cho chúng có màu xanh lục. Thực vật có phôi nói chung có không bào trung tâm phình to, tạo ra sức trương tế bào và giữ cho thực vật được cứng. Chúng không có các roi và trung thể, ngoại trừ ở một số giao tử nhất định.
Tảo là một nhóm lớn và đa dạng, bao gồm các sinh vật thông thường là tự dưỡng, gồm một hay nhiều tế bào có cấu tạo đơn giản, có màu khác nhau, luôn luôn có chất diệp lục nhưng chưa có rễ, thân, lá. Hầu hết tảo sống trong nước. Đây là những sinh vật mà vách thân chứa xenluloza, là những sinh vật tự dưỡng vì chứa diệp lục, quang hợp nhờ ánh sáng và CO2. Cơ quan dinh dưỡng còn gọi là tản. Tảo có nhiều dạng: đơn bào, sợi xiên, sợi phân nhánh, hình ống, hình phiến. Tảo không có mô dẫn truyền.
Phân loại tảo Tảo lục
Tảo lục là một nhóm lớn các loài tảo, mà thực vật có phôi (Embryophyta) (hay thực vật bậc cao) đã phát sinh ra từ đó[1]. Như vậy, chúng tạo nên một nhóm cận ngành, mặc dù nhóm bao gồm cả tảo lục và phân giới Thực vật có phôi là đơn ngành (và thường được biết đến với tên gọi là giới Thực vật - Plantae). Tảo lục bao gồm trùng roi đơn bào và tập đoàn trùng roi (thường nhưng không phải luôn luôn với 2 roi trên 1 tế bào), cũng như các dạng khuẩn cầu và khuẩn sợi, sống thành tập đoàn khác và các dạng tảo biển vĩ mô. Trong bộ Luân tảo (Charales) (quan hệ gần nhất với thực vật đa bào), có sự phân biệt đầy đủ của các mô. Có khoảng 6.000 loài tảo lục[2]. Nhiều loài sống cả đời ở dạng đơn bào, trong khi những loài khác tạo thành dạng tập đoàn, tập đoàn định số (coenobium) hoặc sợi dài hay tảo biển vĩ mô phân dị cao.
Có một vài nhóm sinh vật dựa vào tảo lục để thực hiện chức năng quang hợp của chúng. Lục lạp trong trùng roi xanh (Euglenoidea) và tảo lục phức tạp (Chlorarachnea) là thu được từ việc tiêu hóa thực bào tảo lục[1][3], và ở nhóm thứ hai thì một nhân dấu vết còn lưu lại (hình thái nhân). Tảo lục cũng được tìm thấy là sống cộng sinh trong trùng lông Paramecium, và trong loài thủy tức Hydra viridiscũng như trong một số loài giun dẹt (Platyhelminthes). Vài loài tảo lục, đặc biệt là các chi Trebouxia và Pseudotrebouxia (lớp Trebouxiophyceae), có thể được tìm thấy trong dạng cộng sinh với nấm thành địa y. Nói chung các loại nấm trong địa y không thể sống độc lập, trong khi tảo trong tự nhiên thường không sống với nấm. Các loài tảo lục thuộc chi Trentepohlia sống ký sinh trên vỏ của một số loài cây gỗ.
Tảo Đỏ
Tảo đỏ là những sinh vật quang tự dưỡng thuộc ngành Rhodophyta. Phần lớn các loài rong đều thuộc nhóm này. Các thành viên trong ngành có đặc điểm chung là màu đỏ tươi hoặc tía. Màu sắc của chúng là do các hạt sắc tố phycobilin tạo thành. Phycobilin là sắc tố đặc trưng cho tảo đỏ và vi khuẩn lam. Người ta cho rằng lục lạp của tảo đỏ có nguồn gốc từ vi khuẩn lam cộng sinh với tảo mà thành.
Hiện nay đã phân loại được gần 4.000 loài tảo đỏ, phần lớn sống ở biển, chỉ có một số ít sống ở nước ngọt. Mặc dù tảo đỏ có mặt ở tất cả các đại dương nhưng chúng chỉ phổ biến ở các vùng biển ấm nhiệt đới nơi chúng có thể phân bố sâu hơn bất kỳ một sinh vật quang hợp nào. Tảo đỏ là các sinh vật đa bào và cơ thể phân nhiều nhánh. Tuy nhiên, cơ thể chúng lại không có sự biệt hóa thành các môriêng biệt. Thành tế bào tảo đỏ có một lớp cứng bằng cellulose ở bên trong và một lớp gelatin ở bên ngoài. Tế bào của chúng có thể có một hay nhiều nhân tùy thuộc vào từng loài. Tế bào phân chia bằng cách nguyên phân. Tảo đỏ hoàn toàn không có roi bơi; không có các tế bào có khả năng di chuyển ở bất kỳ dạng nào.
Lạp lục trong tế bào tảo đỏ có phycobilin, chlorophyl a, carotene và xanthophyll. Ở vùng sâu đại dương, ánh sáng xâm nhập tới có bước sóng rất khác so với các thủy vực nông, trong điều kiện đó phycobilin có khả năng hấp thụ ánh sáng tốt hơn so với chlorophyl a. Điều này đã giải thích tại sao tảo đỏ có thể phân bố tới độ sâu 268 m (879 ft). Hợp chất carbonhydrate tích lũy trong tảo đỏ dưới dạng tinh bột floridean, một dạng polymer đặc biệt của glucose khác với dạng tinh bộtcủa các loài thực vật khác.
Chu trình sống của tảo đỏ vô cùng phức tạp, liên quan tới một pha đơn bội và hai pha lưỡng bội. Phần lớn tảo đỏ nước mặn có cơ thể mềm mại, mỏng manh còn được gọi là thalli. Tuy nhiên tảo rạn san hô (coralline algae) có cơ thể được calci hóa nên khá vững chắc. Nó là một phần quan trọng trong việc tạo thành rạn san hô ở các vùng biển nhiệt đới. Vì cấu trúc thành tế bào vững trắc như vậy nên hóa thạch của chúng từ cách đây khoảng 700 triệu năm vẫn còn khá nhiều. Ngày nay người ta có thể chiết suất agar từ một vài giống tảo đỏ để làm môi trường nuôi cấy vi khuẩn và nhiều sinh vật khác. Bên cạnh đó nó cũng là một nguồn iode quan trọng.
Tảo xoắn (tảo nước ngọt)
Tảo xoắn (tên khoa học là Spirulina platensis) là một loại vi tảo dạng sợi xoắn màu xanh lục, chỉ có thể quan sát thấy hình xoắn sợi do nhiều tế bào đơn cấu tạo thành dưới kính hiển vi.Những nghiên cứu mới nhất lại cho biết chúng cũng không phải thuộc chi Spirulina mà lại là thuộc chi Arthrospira. Tên khoa học hiện nay của loài này là Arthrospira platensis, thuộc bộ Oscilatoriales, họ Cyanobacteria.[1] Tảo Spirulina đã được nghiên cứu từ nhiều năm nay. Chúng có những đặc tính ưu việt và giá trị dinh dưỡng cao[2]. Các nhà khoa học trên thế giới đã coi tảo Spirulina là sinh vật có ích cho loài người. Loại tảo này do tiến sĩ Clement người Pháp tình cờ phát hiện vào những năm 1960 khi đến hồ Tchad ở Trung Phi. Nhà khoa học này không khỏi kinh ngạc khi vùng đất cằn cỗi, đói kém quanh năm nhưng những thổ dân ở đây rất cường tráng và khỏe mạnh. Khi Clement tìm hiểu về thức ăn của họ, bà phát hiện trong mùa không săn bắn, họ chỉ dùng một loại bánh màu xanh mà nguyên liệu chính là thứ họ vớt lên từ hồ. Qua phân tích, bà phát hiện ra loại bánh có tên Dihe này chính là tảo Spirulina.
Tổ chức Y tế thế giới (WHO/OMS) công nhận tảo Spirulina là thực phẩm bảo vệ sức khỏe tốt nhất của loài người trong thế kỉ 21. Cơ quan quản lí thực phẩm và dược phẩm Hoa Kì (FDA) công nhận nó là một trong những nguồn protein tốt nhất..
Rong mơ
Rong mơ sống ở nước biển, sống thành từng đám lớn, bám vào đá hoặc san hô nhờ giá bám ở gốc, chưa có rễ, thân, lá. Rong mơ có màu nâu vì trong tế bào ngoài chất diệp lục còn có chất màu phụ màu nâu. Rong mơ cũng quang hợp và tự tạo ra chất dinh dưỡng (dinh dưỡng tự dưỡng). Ngoài sinh sản vô tính, rong mơ còn sinh sản hữu tính (kết hợp giữa tinh trùng và noãn cầu).
Một số tảo khác
Tảo đơn bào gồm tảo tiểu cầu và tảo silic. Tảo đa bào gồm tảo vòng, rau diếp biển, rau câu và tảo sừng hươu.
Vai trò của tảo
Cùng với các thực vật ở nước khác, khi quang hợp, tảo thải ra khí ôxy giúp cho sự hô hấp của các động vật ở nước. Những tảo nhỏ sống trôi nổi là nguồn thức ăn của cá và nhiều động vật ở nước khác. Tảo có thể dùng làm thức ăn cho người và gia súc, ví dụ: tảo tiểu cầu (có nhiều chất dạm và một ít vitamin C, B12), rau câu,... Một số tảo được dùng làm phân bón, làm thuốc, nguyên liệu dùng trong công nghiệp như làm giấy, hồ dán, thuốc nhuộm,...
Tảo cũng có thể gây hại: một số tảo đơn bào sinh sản quá nhanh gây hiện tượng "nước nở hoa", khi chết làm cho nước bị nhiễm bẩn làm chết cá; tảo xoắn, tảo vòng khi sống ở ruộng lúa nước có thể quấn lấy gốc cây làm cây lúa khó đẻ nhánh.
Thực vật học (từ tiếng Hy Lạp cổ đại βοτάνη botane, "đồng cỏ, cỏ, và từ tiếng βόσκειν boskein, "chăn nuôi"[1][2][3]) là một môn khoa học nghiên cứu về thực vật. Thông thường, môn khoa học này bao gồm việc nghiên cứu các loài nấm, tảo và virus. Những người nghiên cứu về thực vật học được gọi là nhà thực vật học. [1][2][3] Thực vật học bao hàm nhiều lĩnh vực nghiên cứu về thực vật như: sinh trưởng, sinh sản, trao đổi chất, phát sinh hình thái (morphogenesis development), bệnh học thực vật (phytopathology), tiến hóa, Phân loại thực vật, sinh lý thực vật, hình thái thực vật, sinh hóa thực vật và hóa thạch thực vật. Ngày này, các nhà thực vật học nghiên cứu khoảng 400000 loài sinh vật sống,[4] trong đó có 260.000 loài thực vật có mạch và khoảng 248.000 là thực vật có hoa.[5]
Sự bắt đầu của các hệ thống phân loại hiện đại có thể lần theo vào thời gian khoảng thế kỷ 15 và 16 khi một số nỗ lực khoa học về phân loại thực vật. Vào thế 19 và 20, nhiều kỹ thuật chủ yếu mới được phát triển để nghiên cứu thực vật, bao gồm kính hiển vi, tính toán nhiễm sắc thể, phân tích hóa học thực vật. Trong 2 thập niên cuối thế kỷ 20, ADN được dùng để phân loại thực vật một cách chính xác hơn.
Lịch sử Thời kỳ đầu
Thực vật học có nguồn gốc từ thảo dược học, việc nghiên cứu và sử dụng thực vật là do các đặc điểm y học của nó.[6] Lịch sử về thực vật học được ghi nhận trước đây bao gồm nhiền tài liệu cổ và các phân loại thực vật. Ví dụ các công trình về thực vật học trước đây được tìm thấy trong các văn bản thánh cổ ở Ấn Độ có tuổi trước 1100 TCN,[7][8] các tài liệu bằng tiếng Avesta cổ,[9] và các công trình ở Trung Quốc trước khi đất nước này được thống nhất năm 221 TCN.[7][10]
Thực vật học hiện đại bắt nguồn từ Hy Lạp Cổ đại, đặc biệt từ Theophrastus (c. 371–287 BC), một học trò của Aristotle ông đã phát minh và miêu tả nhiều nguyên lý của thực vật học và được cộng đồng khoa học xem là "cha đẻ của Thực vật học".[11] Các công trình chính của ông Historia Plantarum (Khảo sát hực vật) và On the Causes of Plants, chứa nhiều đóng góp quan trọng nhất cho khoa học thực vật cho đến thời kỳ Trung Cổ, gần 17 thế kỷ sau khi chúng được viết ra.[11][12]
Một công trình khác thời Hy Lạp Cổ đại cũng ảnh hưởng đến thực vật học thời kỳ này là De Materia Medica, một bách khoa toàn tư có 5 quyển về thảo dược được viết vào giữa của thế kỷ thứ nhất của một nhà dược học và thầy thuốc Hy Lạp Pedanius Dioscorides. De Materia Medica được tiếp nhận trong hơn 1500 năm.[13] Các đóng góp quan trọng từ thế giới Hồi giáo trung cổ như Nabatean Agriculture của Ibn Wahshiyya, Book of Plants của Abū Ḥanīfa Dīnawarī (828–896), và The Classification of Soils của Ibn Bassal. Vào đầu thế kỷ 13, Abu al-Abbas al-Nabati, và Ibn al-Baitar (d. 1248) đã viết về thực vật học một cách hệ thống và khoa học.[14][15][16]
Vào giữa thế kỷ 16, "botanical gardens" được xây dựng nhiều ở các trường đại học Ý – Padua botanical garden năm 1545 thường được xem là vườn đầu tiên vẫn còn tọa lạc tại vị trí ban đầu của nó. Các khu vườn này vẫn là nơi thực hành của các "physic gardens" trước đây, các loài cây ở đây được dùng vào mục đích y học. Chúng giúp phát triển ngành thực vật học ở góc độ đối tượng nghiên cứu hàn lâm. Các bài giảng về sự phát triển của thực vật trong các khu vườn và tác dụng y học của chúng đã được minh họa. Các vườn thực vật hình thành muộn hơn ở ở miền bắc châu Âu; vườn đầu tiên ở Anh là vườn thực vật Đại học Oxford năm 1621. Trong suốt thời gian này, thực vật học vẫn phụ thuộc nhiều vào y học.[17]
Bác sĩ người Đức Leonhart Fuchs (1501–1566) là một trong "3 cha đẻ người Đức của thực vật học", cùng với nhà thần học Otto Brunfels (1489–1534) và bác sĩ Hieronymus Bock (1498–1554).[18][19] Fuchs và Brunfels đã tách ra khỏi việc sao chép các công trình truyền thống trước kia để tạo ra các nghiên cứu gốc của riêng họ. Bock đã tạo ra một hệ thống phân loại riêng của ông.
Bác sĩ Valerius Cordus (1515–1544) là tác giả quyển sách về thảo dược quan trọng về dược liệu và thực vật Historia Plantarum năm 1544 và tầm quan trọng lâu dài của dược điển Dispensatorium năm 1546.[20] Nhà tự nhiên học Conrad von Gesner (1516–1565) và thảo dược học John Gerard (1545–c. 1611) đã công bố các thảo dược bao gồm các loài việc sử dụng các loài thực vật. Nhà tự nhiên học Ulisse Aldrovandi (1522–1605) được xem là cha đẻ của lịch sử tự nhiên, bao gồm việc nghiên cứu về thực vật. Năm 1665, việc sử dụng kính hiển vi đầu tiên, nhà bác học Robert Hooke đã phát minh ra các tế bào trong một cái nút chai, và một thời gian ngắn sau đó trong các mô thực vật sống.[21]
Thực vật học cận đại
Vườn Linnaeus, nơi ở của Linnaeus ở Uppsala, Thụy Điển, được trồng theo Systema sexuale của ông
Trong thế kỷ 18, hệ thống định danh thực vật được phát triển tương đương với khóa lưỡng phân, theo đó các thực vật chưa được định danh được xếp vào các nhóm phân loại học (như họ, chi và loài) bằng cách lập một loạt các lựa chọn giữa các cặp tính trạng. Việc lựa chọn và trình tự của các tính trạng có thể nhân tạo trong các khóa được thiết kế một cách hoàn toàn cho việc định danh hoặc liên quan chặt chẽ với bậc ngành của phân loại học trong các khóa tổng quát.[22] Cho đến thế kỷ 18, các thực vật mới trong nghiên cứu đã đến châu Âu với số lượng ngày càng tăng từ các nước được phát hiện mới và các quốc gia thuộc địa của châu Âu trên toàn thế giới. Năm 1753 Carl von Linné (Carl Linnaeus) đã xuất bản quyển Species Plantarum, bao gồm việc phân loại theo thứ bậc các loài thực vật mà hiện vẫn còn là tham chiếu trong danh mục thực vật hiện đại. Sự kiện này đã thiết lập nên một cơ chế đặt tên hai phần hay danh pháp hai phần chuẩn hóa gồm phần đầu là tên chi và phần sau là tên loài trong chi đó.[23] Đối với mục đích phân loại, Systema Sexuale của Linnaeus đã phân thực vật thành 24 nhóm theo số lượng các cơ quan sinh sản đực của chúng. Nhóm thứ 24, Cryptogamia, bao gồm tất cả các loài thực vật có bộ phân sinh sản ẩn, rêu, địa tiền (liverwort), dương xỉ, tảo và nấm.[24]
Khi hiểu biết về giải phẫu, hình thái và vòng đời thực vật tăng lên người ta đã nhận ra rằng có sự tương đồng tự nhiên giữa thực vật hơn hệ thống giới tính không tự nhiên mà Linnaeus đã chỉ ra. Adanson (1763), de Jussieu (1789), và Candolle (1819) đều đề xuất nhiều hệ thống phân loại tự nhiên thay thế theo đó các thực vật được gộp lại dựa trên các đặc điểm giống nhau và được chấp nhận rộng rãi. Hệ thống Candolle phản ánh những tư tưởng của ông về sự tiến triển phức tạp của hình thái và phân loại sau đó của Bentham và Hooker chịu ảnh hưởng theo cách tiếp cận của Candolle. Ấn phẩm của Darwin,Về nguồn gốc các loài năm 1859 và quan điểm có chung nguồn gốc của ông cần phải có những sửa đổi đối với hệ thống Candolle để phản ánh các mối quan hệ tiến hóa như là sự khác biệt về tương đồng chỉ về mặt hình thái.
thực vật học được thúc đẩ rất nhiều bởi sự xuất hiện của quyển sách "hiện đại" đầu tiên của Matthias Schleiden Grundzüge der Wissenschaftlichen Botanik, được xuất bản đầu tiên bằng tiếng Anh năm 1849 với tựa đề Principles of Scientific Botany.[25] Schleiden là một nhà kính hiển vi học và giải phẫu thực vật, ông là đồng sáng lập nên học thuyết tế bào cùng với Theodor Schwann và Rudolf Virchow và là một trong những người đầu tiên hiểu được ý nghĩa của nhân tế bào do Robert Brown miêu tả năm 1831.[26] Năm 1855, Adolf Fick thành lập Fick's laws cho phép tính toán tốc độ khuếch tán phân tử trong các hệ sinh học.
Hóa thạch là những di tích và di thể (xác) của sinh vật được bảo tồn trong các lớp đá, là đối tượng nghiên cứu chủ yếu của của ngành sinh vật học... Trải qua một thời gian dài tồn tại của Trái đất, đã có rất nhiều sinh vật từng sinh sống. Những sinh vật này sau khi chết, xác hoặc những dấu vết của các hoạt động sống là những chứng cứ được lưu giữ lại, theo sau một thời gian xác của chúng bị phân hủy (thối rữa), chỉ những bộ phần cứng như vỏ xương hoặc cành cây... được bao bọc bởi các trầm tích vật và trải qua quá trình hóa thạch để trở thành đá, nhưng vẫn giữ lại được những hình thái kết cấu (thậm chí một vài chi tiết nhỏ cấu tạo bên trong) đồng thời những dấu vết hoạt động của những sinh vật thời kỳ đó cũng được bảo lưu như vậy.
Nguồn gốc tên gọi "hóa thạch" trong tiếng Anh, từ "fossil" bắt nguồn từ latinh "fossilis" với nghĩa là "đào lên".
Lịch sử nghiên cứu hóa thạch
Trong những ghi chép còn để lại từ xa xưa, có một nhóm học giả người Hy Lạp đã rất ngạc nhiên trước những di tích tồn tại của cá, vỏ sò và một vài dạng sinh vật biển được tìm thấy ở trên núi và sa mạc. Năm 450 trước công nguyên Herodotus đã đề cập tới sa mạc Ai Cập và cũng khẳng định rằng ở đó trước đây một phần bị bao phủ bởi biển Địa Trung Hải, năm 400 trước công nguyên Aristoteles tuyên bố: "hóa thạch là do vật chất hữu cơ tạo thành, nhưng hóa thạch bị ép vào trong tầng nham thạch là do một tác dụng làm mềm trong vỏ trái đất gây ra". Một học trò của ông là Theophrastus năm 350 trước công nguyên cũng đã đưa ra được một vài hóa thạch của các dạng sinh vật, nhưng lại cho rằng hóa thạch do trứng và hạt của cây trong lớp nham phát triển mà thành....
Strabo (63-20 trước công nguyên) cũng chú ý đến những hóa thạch của các sinh vật biển tìm thấy dưới lớp đất dưới đáy.
Trong gia đoạn đen tối của thời kỳ trung cổ, con người có nhiều cách lý giải khác nhau về hóa thạch. Có người cho rằng đó là hiện tượng kỳ bí của thiên nhiên hoặc đó là sản phẩm của ma quỷ tạo ra để mê hoặc con người. Những lời đồn đại mê tín và sự cấm đoán của các tôn giáo đã cản trở việc nghiên cứu hóa thạch vài trăm năm. Mãi đến đầu thế kỷ 15 là thời kỳ khởi nguồn cho việc phổ biến và tiếp thu những kiến thức về hóa thạch, con người đã hiểu rằng hóa thạch là tàn tích của những sinh vật trước kia, nhưng vẫn cho rằng đó là những dấu vết của cuộc đại hồng thủy được ghi trong thánh kinh. Các nhà khoa học và các nhà thần học đã tranh cãi nhau suốt 300 năm sau đó. Đến thời kỳ phục hưng có một vài nhà khoa học đầu tiên mà tiêu biểu là Leonardo da Vinci (1452-1519) đã đề cập tới hóa thạch và đã kiên quyết phủ nhận việc liên quan của hóa thạch với đại hồng thủy, và cũng đã phủ nhận ý kiến cho rằng hóa thạch chỉ xuất hiện ở trên núi cao, họ cũng tin rằng hóa thạch là chứng cứ của những sinh vật cổ đại và cũng cho rằng
Biển Địa Trung Hải trước đây đã bao phủ toàn bộ nước Ý trước đây khi đáy biển lên xuống do các hoạt động địa chất hình thành lên bán đảo Italy.
Xác của sinh vật cổ đại vẫn được lưu giữ lại trong bề mặt đáy biển.
Cuối thế kỷ 18 và đầu thế kỷ 19 đã hình thành lên được một cơ sở vững chắc cho việc nghiên cứu hóa thạch và tạo lên một môn khoa học. Từ đó đến nay hóa thạch thường phát hiện chủ yếu tại tầng nham trầm tích ngoài biển. Khi các trầm tích vật như đá vôi, vụn cát, vỏ xương của động vật bị đè nén rồi kết dính với nhau tạo thành nham, và tạo lên một phần nham trầm tích mặt biển (seafacies). Rất hiếm gặp hóa thạch xuất hiện trong các nham núi lửa và nham biến chất (metamorphic rock) bởi vì:
Nham núi lửa ở trạng thái trước đó là dạng nóng chảy và không tồn tại sự sống.
Còn Nham biến chất đã trải qua những biến đổi rất lớn mà thành khiến cho hình dạng ban đầu của hóa thạch trong đó hầu như bị phá hủy và không còn lại nguyên vẹn. Tuy vậy nếu như may mắn còn được giữ lại trong lớp nham trầm tích một vài hóa thạch thì đó chỉ là một bộ phận rất nhỏ động thực vật thuộc thời kỳ đó mà thôi.
Bằng việc kiểm chứng những điều kiện khắc nghiệt trong quá trình hình thành lên hóa thạch cũng dễ hiểu vì sao những vết tích còn lưu lại trong lớp nham trầm tích (sedimentary rock) cũng chỉ là một bộ phận rất nhỏ còn sót lại từ những sinh vật thuộc thời kỳ trước.
Điều kiện hình thành hóa thạch
Mặc dù một sinh vật có thể hình thành lên hóa thạch hay không đều được quyết định bởi rất nhiều những nhân tố, nhưng có 3 nhân tố cơ bản nhất:
Sinh vật nhất thiết phải có những bộ phận cứng khó phân hủy như xương, vỏ răng và gỗ.... sau đó ở vào một điều kiện vô cùng thuận lợi cho dù là những sinh vật mềm yếu như côn trùng hoặc sứa cũng có thể tạo hình thành lên hóa thạch.
Sinh vật khi chết phải được bảo vệ để tránh khỏi những tác động phá hủy, nếu như các phần cơ thể của nó bị nghiền nát hoặc bị ăn mòn thì khả năng tạo thành hóa thạch của sinh vật không thể thực hiện được.
Sinh vật cần thiết phải được chôn xuống và bao phủ bởi những vật chất có thể giúp nó chống lại những điều kiện khắc nghiệt có thể khiến nó bị phân rã. Xác của những sinh vật ở biển thường rất dễ hình thành hóa thạch. Bởi vì xác sinh vật biển chết sau khi lắng xuống đáy biển bị cát phủ lấp, cát trong những niên đại địa chất sau đó sẽ biến thành đá vôi (limestone) hoặc diệp thạch (schist), những trầm tích vật (sediment) nhỏ không dễ làm tổn hại đến xác của sinh vật.
Giới Nấm (tên khoa học: Fungi) bao gồm những sinh vật nhân chuẩn tự dưỡng có thành tế bào bằng kitin (chitin). Phần lớn nấm phát triển dưới dang các sợi đa bào được gọi là sợi nấm (hyphae) tạo nên hệ sợi (mycelium), một số nấm khác lại phát triển dưới dạng đơn bào. Quá trình sinh sản (hữu tính hoặc vô tính) của nấm thường qua bào tử, được tạo ra trên những cấu trúc đặc biệt hay thể quả. Một số loài lại mất khả năng tạo nên những cấu trúc sinh sản đặc biêt và nhân lên qua hình thức sinh sản sinh dưỡng.
Những đại diện tiêu biểu của nấm là nấm mốc, nấm men và nấm lớn (nấm quả thể). Giới Nấm là nhóm sinh vật đơn ngành (monophyletic) mà có nguồn gốc hoàn toàn khác biệt với những sinh vật có hình thái tương tự như nấm nhầy (myxomycetes) hay mốc nước (oomycetes). Nấm có mối quan hệ gần với động vật hơn thực vật, cho dù thế thì môn học về nấm, hay nấm học, lại thường được xếp vào thành một nhánh của thực vật học.
Trên Trái Đất, đa phần các nấm đều không thể nhìn thấy được bằng mắt thường, chúng sống phần lớn ở trong đất, chất mùn, xác sinh vật chết, cộng sinh hoặc kí sinh trên cơ thể động, thực vật và nấm khác. Vi nấm đóng một vai trò quan trọng trong hệ sinh thái, chúng phân hủy các vật chất hữu cơ và không thể thiếu được trong chu trình chuyển hóa và trao đổi vật chất. Một số loài nấm có thể nhận thấy được khi ở dạng thể quả, như nấm lớn và nấm mốc. Nấm được ứng dụng rất rộng rãi trong đời sống lẫn sản xuất, nhiều loài được sử dụng trong công nghệ thực phẩm, sử dụng làm thức ăn hoặc trong quá trình lên men. Nấm còn được dùng để sản xuất chất kháng sinh, hoóc môn trong y học và nhiều loại enzym. Tuy vậy, nhiều loại nấm lại có chứa các chất hoạt động sinh học được gọi là mycotoxin, như ancaloit và polyketit, là những chất độc đối với động vật lẫn con người. Một số loại nấm được sử dụng để kích thích hoặc trong các nghi lễ truyền thống với vai trò tác động lên trí tuệ và hành vi của con người. Vài loại nấm có thể gây ra các chứng bệnh cho con người và động vật, cũng như bệnh dịch cho cây trồng, mùa màng và có thể gây tác động lớn lên an ninh lương thực và kinh tế.
Sự đa dạng
Nấm phân bố trên toàn thế giới và phát triển ở nhiều dạng môi trường sống khác nhau, kể cả sa mạc, nơi tập trung nồng độ muối cao[3] hay có phóng xạ ion hóa,[4] cũng như trầm tích biển sâu.[5] Đa phần nấm sống ở trên cạn, nhưng một số loài sống ở môi trường nước (như Batrachochytrium dendrobatidis - ký sinh và làm suy giảm số lượng động vật lưỡng cư toàn cầu). Nấm thủy sinh còn sống ở vùng nhiệt dịch đại dương.[6] Nấm và vi khuẩn là những sinh vật phân huỷ chính có vai trò quan trọng đối với các hệ sinh thái trên cạn trên toàn thế giới. Dựa theo sự theo tỉ lệ giữa số loài nấm với số loài thực vật ở trong cùng một môi trường, người ta ước tính giới Nấm có khoảng 1,5 triệu loài[7]. Khoảng 100.000 loài nấm đã được các nhà phân loại học phát hiện và miêu tả,[8] tuy nhiên kích cỡ thực sự của tính đa dạng của giới Nấm vẫn còn là điều bí ẩn[9]. Đa phần nấm phát triển dưới dạng các sợi đa bào gọi là sợi nấm, cấu tạo nên thể sợi (hay khuẩn ty), trong khi những loài khác thì lại phát triển dưới dạng đơn bào[10][11]. Cho đến gần đây, nhiều loại nấm đã được miêu tả dựa trên những đặc điểm hình thái, như kích cỡ và hình dạng các bào tử hay thể quả, hay dựa trên khái niệm loài sinh vật với sự trợ giúp của các công cụ phân tử, như phương pháp Dideoxy, đã gia tăng mạnh cách thức và khả năng ước tính sự đa dạng của nấm trong phạm vi các nhóm phân loại khác nhau[12].
Hình thái
Vòng đời
Sinh thái
Nấm lỗ phát triển trên một cây ở Borneo
Dù không dễ thấy, nhưng nấm lại có mặt ở tất cả các môi trường trên Trái Đất và đóng một vai trò rất quan trọng trong hệ sinh thái. Cùng với vi khuẩn, nấm là sinh vật phân hủy chính ở hầu hết các hệ sinh thái trên cạn (và có thể dưới nước), bởi vậy nên chúng cũng có vai trò quan trọng các chu trình sinh địa hóa và ở nhiều lưới thức ăn. Khi sống hoại sinh hay cộng sinh, chúng phân hủy những vật chất hữu cơ thành những phân tử vô cơ, rồi sau đó những chất này sẽ được đồng hóa ở thực vật hay những sinh vật khác
Cộng sinh
Nấm có mối quan hệ cộng sinh với hầu hết tất cả các giới [15][16][17]. Quan hệ của chúng có thể hỗ trợ hoặc đối nghịch nhau, hay với những nấm hội sinh thì không đem lại bất cứ lợi ích hay tác hại rõ ràng nào đối với vật chủ [18][19][20].
Với thực vật
Một nấm rễ thạch nam tách từ Woollsia pungens
Nấm rễ là một hình thức cộng sinh giữa thực vật và nấm, chia làm hai loại: nấm rễ trong (endomycorrhiza, tức nấm kí sinh đơn bào sống bên trong tế bào rễ cây) và nấm rễ ngoài (ectomycorrhiza, tức rễ của nấm bám dày đặc xung quanh đầu rễ cây và xâm nhập vào giữa các tế bào rễ cây). Đây là quần hợp nấm-thực vật được biết nhiều nhất và đóng vai trò quan trọng trong quá trình phát triển của thực vật cũng như nhiều hệ sinh thái, hơn 90% các loài thực vật có quan hệ với nấm theo hình thức nấm rễ và phụ thuộc vào mối quan hệ này để tồn tại[21][22][23]. Sự cộng sinh nấm rễ đã có lịch sử xa xưa, ít nhất là từ hơn 400 triệu năm về trước [24]. Chúng thường làm tăng khả năng hấp thu các hợp chất vô cơ của thực vật, như nitrat và photphat, từ những đất có nồng độ những nguyên tố thiết yếu thấp[25]. Ở một số nấm rễ, thành phần nấm có thể đóng vai trò trung gian giữa thực vật với thực vật, vận chuyển carbohydrate và các chất dinh dưỡng khác. Những cộng đồng nấm rễ đó được gọi là "mạng lưới nấm rễ chung"[26]. Một số nấm có khả năng kích thích sự sinh trưởng của cây bằng cách tiết ra các hoóc môn thực vật như axít idolaxetic (IAA) [27].
Địa y là dạng cộng sinh giữa nấm (hầu hết các loài nấm nang và một số nấm đảm) với tảo hay vi khuẩn lam (gọi chung là đối tác quang hợp), trong đó những tế bào quang hợp được gắn vào những mô nấm [28]. Giống với nấm rễ, những đối tác quang hợp sẽ cung cấp cacbohyđrat được tạo ra trong quá trình quang hợp, đổi lại nấm cung cấp cho chúng các chất khoáng và nước. Những chức năng của toàn bộ cơ thể địa y gần như giống hệt với một cơ thể đơn độc. Địa y là những sinh vât tiên phong và xuất hiện ở những nơi nguyên thủy như đá tảng hay nham thạch núi lửa đã nguội. Chúng có thể thích nghi cực tốt với những điều kiện khắc nghiệt như giá lạnh hay khô hạn và là những ví dụ tiêu biểu nhất của sự cộng sinh[27].
Một số loài nấm sống trong cây có thể tiết ra những độc tố nấm để ngăn cản những động vật ăn cỏ ăn vật chủ của chúng[29].
Với côn trùng
Nhiều côn trùng có mối quan hệ hỗ trợ với nhiều loại nấm. Vài loại kiến trồng những loài nấm thuộc bộ Nấm mỡ (Agaricales) để làm nguồn thức ăn chính, trong khi đó những loài bọ cánh cứng Ambrosia trồng nhiều loài nấm trong lớp vỏ cây mà chúng cư trú [30]. Loài mối ở xavan châu Phi cũng được biết có khả năng trồng nấm [31].
Mầm bệnh và kí sinh
Bệnh đạo ôn ở lúa do Magnaporthe oryzae gây ra
Tuy vậy, nhiều loại nấm lại kí sinh trên thực vật, động vật (cả con người) và nấm khác. Những loài nấm gây bệnh trên cây trồng có thể gây thiệt hại rộng lớn cho ngành nông nghiệp và lâm nghiệp, ví dụ như nấm đạo ôn (Magnaporthe oryzae) gây bệnh cho lúa[32], Ophiostoma ulmi và Ophiostoma novo-ulmi gây ra bệnh du Hà Lan[33], còn Cryphonectria parasitica là nguyên nhân của bệnh thối cây dẻ[34]. Những loài gây bệnh cho cây thuộc các chi Fusarium, Ustilago, Alternaria và Cochliobolus[19], còn những loài có khả năng gây bệnh cho người lại thuộc các chi như Aspergillus, Candida, Cryptoccocus[20][35][36], Histoplasma[37] và Pneumocystis [38]. Chúng có thể gây ra những bệnh ngoài da ở người như nấm chân hay hắc lào cho đến những bệnh nguy hiểm có thể gây chết người như viêm màng não (nấm Cryptococcus neoformans)[39] hay viêm phổi. Nấm gây ra nhiều bệnh cơ hội, tức những bệnh tấn công những người bị suy giảm miễn dịch [40], trong đó có những người bị HIV/AIDS [41], ví dụ như bệnh candidiasis (nấm Candida, gây ra chứng lở miệng ở trẻ em và âm đạo phụ nữ), histoplasmosis (Histoplasma capsulatum), cryptococcosis (Cryptococcus neoformans), aspergillosis (Aspergillus), coccidioidomycosis (Coccidioides immitis hay C. posadasii), viêm phổi pneumocystis (Pneumocystis jirovecii)... và rất nhiều bệnh khác[41][42].
Có khoảng 70 loài[43] nấm sinh bào tử là những tác nhân gây dị ứng. Chúng có thể là nấm mốc trong nhà hay ngoài trời, đa phần là nấm sợi như các chi Alternaria, Aspergillus, Cladosporium, Helminthosporium, Epicoccum, Penicillium, Fusarium..., chỉ có vài loài là nấm đơn bào như Candida, Rhodotorula, có một số loài là nấm lớn như Agaricus, Coprinus, Fomes, Ganoderma...[43][44] Bào tử nấm có thể gây ra những chứng như hen suyễn, viêm mũi dị ứng[45], các bệnh nấm dị ứng phế quản phổi và viêm phổi quá mẫn[44].
Săn mồi
Một số loài nấm là những kẻ săn giun tròn. Chúng có thể biến đổi sợi nấm để tạo thành những cấu trúc đặc biệt có chức năng bẫy giun tròn, nên được gọi với tên chung là nấm bẫy mồi[46]}. Những loại bẫy thường thấy là: mạng dính hay lưới dính, bọng dính, vòng không thắt, cột dính, vòng thắt[47][48] và bào tử dính[49]. Các loài nấm bắt mồi theo kiểu này thường thuộc các chi Arthrobotrys, Dactylaria, Dactylella và Trichothecium[46][50]. Có vài loài như Zoopage phanera[46][51] thì lại tiết chất dính ra toàn bộ mặt ngoài sợi nấm và cũng có khả năng bẫy mồi tương tự.
Dinh dưỡng và khả năng tự dưỡng
Sự phát triển của nấm dưới dạng sợi nấm ở những môi trường rắn cũng như dưới dạng đơn bào ở môi trường nước, đều được điều chỉnh để hút các chất dinh dưỡng hiệu quả nhất từ môi trường, bởi chúng đều có tỉ lệ diện tích bề mặt trên thể tích cao. Sự thích nghi hình thái đã được bổ sung bởi những enzym thủy phân trong những môi trường tiêu hóa có phân tử hữu cơ lớn, như polysaccarit, protein, lipit và những chất nền dinh dưỡng khác. Những phân tử này bị thủy phân thành những phân tử nhỏ hơn[52][53][54], sau đó trở thành những chất dinh dưỡng được hấp thu vào tế bào nấm.
Thông thường nấm được coi là những sinh vật dị dưỡng, tức những cơ thể chỉ có thể lấy cacbon từ những sinh vật khác cho quá trình trao đổi chất, tuy nhiên nấm đã tiến hóa khả năng chuyển hoá mà cho phép chúng sử dụng đa dạng những loại chất nền hữu cơ để phát triển, bao gồm các hợp chất đơn giản như nitrat, amoniac, axetat hay êtanol[55][56]. Những nghiên cứu gần đây cho thấy khả năng rằng nấm đã sử dụng sắc tố melanin để lấy năng lượng từ những phóng xạ ion hóa, như tia gamma, gọi nôm na là "vô tuyến dưỡng"[57]. Người ta cho rằng quá trình này có điểm tương đồng với quá trình quang hợp ở thực vật[57], tuy nhiên hiện nay đang thiếu những bằng chứng sinh hóa có giá trị ủng hộ cho giả thuyết này.
Vai trò đối với con người
Nấm đã được con người sử dụng để chế biến và bảo quản thức ăn một cách rộng rãi và lâu dài: nấm men được sử dụng cho quá trình lên men để tạo ra rượu, bia và bánh mì, một số loài nấm khác được sử dụng để sản xuất xì dầu và tempeh. Trồng nầm và hái nấm là những ngành kinh doanh lớn ở nhiều nước. Nhiều loại nấm được sử dụng để sản xuất chất kháng sinh, gồm các kháng sinh β-lactam như penicillin và cephalosporin. Những loại kháng sinh này đều được sử dụng rộng rãi trong việc chữa trị các bệnh do vi khuẩn, như lao, phong cùi, giang mai và nhiều bệnh khác ở đầu thế kỷ 20 và tiếp tục đóng một vai trò quan trọng trong hóa học trị liệu kháng khuẩn. Môn khoa học nghiên cứu về lịch sử ứng dụng và vai trò của nấm được gọi là nấm học dân tộc.
Chế biến thực phẩm
Nấm men khô, dạng kết tinh của men được sử dụng thương mại
Bình Kombucha đang lên men
Nấm men bánh mì với loài chính là Saccharomyces cerevisiae, một nấm đơn bào, được sử dụng rộng rãi trong việc làm bánh mì và những sản phấm từ bột mì khác, như pizza hay bánh bao [58]. Trong quá trình lên men rượu dưới điều kiện yếm khí, nấm men sẽ sản sinh ra rượu etanol và khí cacbonic (CO2) có vai trò làm nở bột mì. Một số loài nấm men của chi Saccharomyces cũng được sử dụng để sản xuất đồ uống có cồn, như bia hay rượu vang thông qua quá trình lên men rượu[59]. Nấm men và vi khuẩn acetic được sử dụng trong quá trình chuẩn bị Kombucha, một loại trà ngọt lên men. Những loại men tìm thấy trong trà rất đa dạng, như Brettanomyces bruxellensis, Candida stellata, Schizosaccharomyces pombe, Torulaspora delbrueckii và Zygosaccharomyces bailii[60]. Ngoài ra, Vegemite và Marmite, những chiết xuất nấm men[61], đã được sử dụng rộng rãi làm gia vị để chế biến thức ăn trong đời sống.
Vân xanh pho mát Stilton với nấm Penicillium roqueforti.
Một số loài nấm sợi thường được sử dụng để sản xuất sinh khối protein. Mốc Aspergillus oryzae được sử dụng rộng rãi ở nhiều nước, như sản xuất nước tương (xì dầu), súp miso và rượu sake ở Nhật Bản[62] hay làm tempeh ở Java[63]. Mốc hoa cau được dùng để sản xuất tương, loại thực phẩm phổ biến ở Việt Nam, cũng chính là A. oryzae[62], tuy nhiên tương sản xuất thủ công lại có độ an toàn không cao, bởi những loại mốc tốt và không độc như A. oryzae và Aspergillus sojae lại rất dễ lẫn lộn với những loại mốc nguy hiểm có độc tố gây ung thư khác là A. flavus và A. parasiticus[62]. Quorn là loại thực phẩm giàu protein được sản xuất từ mốc Fusarium venenatum[64], và được dùng trong việc chế biến đồ ăn chay. Nhiều loại thực phẩm khác cũng được chế biến bởi mốc như chao (ủ nhờ mốc Actinomucor elegans, Mucor racemosus hay Rhizopus)[65], ang-kak (gạo lên men với mốc đỏ Monascus purpureus)[66], salami (một loại xúc xích, lên men nhờ P. nalgiovense, P. chrysogenum)[67][68]. Trong sản xuất pho mát, một kinh nghiệm thông thường là cấy bào tử nấm vào sữa đông để tạo mốc, việc này sẽ cho ra hương vị và kết cấu đặc biệt độc nhất của pho mát. Ở những loại pho mát xanh, như Stilton hay Roquefort, thì những vân xanh được tạo ra bởi loài nấm Penicillium roqueforti[69].
Sinh vật đơn bào là các sinh vật mà cơ thể chỉ được cấu tạo từ một tế bào. Tế bào này có thể là sinh vật nhân sơ hoặc sinh vật nhân chuẩn. Một số sinh vật đơn bào có thể hợp tác với nhau để phát triển thành tập đoàn.
Hầu hết các sinh vật nguyên sinh đều là đơn bào.
Tảo bong bóng Valonia ventricosa, một trong những sinh vật đơn bào to nhất với đường kính hơn 1 cm[1]
Paramecium aurelia, một nguyên sinh vật thuộc ngành trùng lông. Hình chụp dưới kính hiển vi quang học
Chlamydomonas reinhardtii, nguyên sinh vật thuộc lớp Tảo lục
Trùng biến hình trần Amoeba proteus, một đại diện tiêu biểu của đơn bào đơn giản
Vi khuẩn (tiếng Anh và tiếng La Tinh là bacterium, số nhiều bacteria) đôi khi còn được gọi là vi trùng, là một nhóm (giới hoặc vực) vi sinh vật nhân sơ đơn bào có kích thước rất nhỏ; một số thuộc loại ký sinh trùng. Vi khuẩn là một nhóm sinh vật đơn bào, có kích thước nhỏ (kích thước hiển vi) và thường có cấu trúc tế bào đơn giản không có nhân, bộ khung tế bào (cytoskeleton) và các bào quan như ty thể và lục lạp. Cấu trúc tế bào của vi khuẩn được miêu tả chi tiết trong mục sinh vật nhân sơ vì vi khuẩn là sinh vật nhân sơ, khác với các sinh vật có cấu trúc tế bào phức tạp hơn gọi là sinh vật nhân chuẩn.
Vi khuẩn là nhóm hiện diện đông đảo nhất trong sinh giới. Chúng hiện diện khắp nơi trong đất, nước, chất thải phóng xạ, suối nước nóng,[2] và ở dạng cộng sinh và kí sinh với các sinh vật khác, và được biết là phát triển mạnh mẽ trong các tàu không gian có người lái.[3] Nhiều tác nhân gây bệnh (pathogen) là vi khuẩn. Hầu hết vi khuẩn có kích thước nhỏ, thường chỉ khoảng 0.5-5.0 μm, mặc dù có loài có đường kính đến 0,3mm (Thiomargarita). Chúng thường có vách tế bào, như ở tế bào thực vật và nấm, nhưng với thành phần cấu tạo rất khác biệt (peptidoglycan). Nhiều vi khuẩn di chuyển bằng tiên mao (flagellum) có cấu trúc khác với tiên mao của các nhóm khác.
Có khoảng 40 triệu tế bào vi khuẩn trong một gram đất và hàng triệu tế bào trong một mm nước ngọt. Ước tính có khoảng 5×1030 vi khuẩn trên Trái Đất,[4] tạo thành một lượng sinh khối vượt hơn tất cả động vật và thực vật.[5] Vi khuẩn có vai trò quan trọng trong tái chế chất dinh dưỡng như cố định nitơ từ khí quyển và gây thối rữa sinh vật khác. Trong vùng dinh dưỡng quanh cách mạch nhiệt dịch và lỗ phun lạnh, vi khuẩn cung cấp những chất dinh dưỡng cần thiết cho sự sống bằng cách biến đổi các hợp chất hòa tan như hydro sulphua và metan thành năng lượng, chúng có thể phát triển mạnh ở nơi sâu nhất trên Trái Đất là rãnh Mariana.[6][7] Các nghiên cứu khác liên quan cũng chỉ ra rằng chúng có thể sống bên trong các đá ở độ sâu 1900 feet bên dưới đáy biển và cách ngoài khơi bờ biển tây bắc Hoa Kỳ 8500.[6][8]
Nguồn gốc và tiến hóa
Tổ tiên của vi khuẩn hiện đại là các sinh vật đơn bào, đó là các dạng sống xuất hiện đầu tiên trên Trái Đất cách nay 4 tỉ năm. Trong vòng 3 tỉ năm, tất cả các sinh vật là các vi sinh vật, và vi khuẩn và vi khuẩn cổ là các dạng sống chủ yếu trên Trái Đất.[9][10] Mặc dù các hóa thạch vi khuẩn đã được tìm thấy như stromatolite, nhưng chúng thiếu các hình thái đặc biệt để xem xét lịch sử tiến hóa của vi khuẩn, hoặc thời điểm xuất phát một loài vi khuẩn đặc biệt. Tuy nhiên, trình tự gen có thể được sử dụng để tái dựng phát sinh loài của vi khuẩn, và các nghiên cứu này chỉ ra rằng vi khuẩn bắt đầu phân nhánh đầu tiên từ dòng vi khuẩn cổ/nhân chuẩn.[11]
Vi khuẩn cũng liên quan đến lần phân nhánh tiến hóa lớn lần thứ 2 của vi khuẩn cổ và nhân chuẩn. Các sinh vật nhân chuẩn là kết quả của sự tham gia của các vi khuẩn trước đó vao tập hợp nội cộng sinh với các tổ tiên của các tế bào nhân chuẩn, mà bản thân chúng có thể liên quan đến vi khuẩn cổ.[12][13] Quá trình này liên quan đến sự nhấn chìm bởi các tế bào nhân chuẩn nguyên thủy của sự cộng sinh alpha-proteobacterial để tạo thành hoặc là mitochondria hoặc là hydrogenosome, chúng vẫn được tìm thấy trong tất cả các sinh vật nhân chuẩn đã được biết đến như trong động vật nguyên sinh cổ "amitochondrial". Sau đó, một số sinh vật nhân chuẩn đã chứa ty thể cũng nhấn chìm các sinh vật giống như cyanobacterial. Điều này dẫn đến sự hình thành lục lạp trong tảo và thực vật. Cũng có một số tảo có nguồn gốc từ thận chí là các sự kiện nội cộng sinh sau đó. Ở đây, eukaryota đã nhấn chìm tảo eukaryotia đã phát triển thành một "thế hệ thứ 2".[14][15] Đây được gọi là sự kiện nội cộng sinh thứ 2.
Lịch sử nghiên cứu và phân loại
Vi khuẩn đầu tiên được quan sát bởi Antony van Leeuwenhoek năm 1683 bằng kính hiển vi một tròng do ông tự thiết kế. Tên "vi khuẩn" được đề nghị sau đó khá lâu bởi Christian Gottfried Ehrenberg vào năm 1828, xuất phát từ chữ βακτηριον trong tiếng Hy Lạp có nghĩa là "cái que nhỏ". Louis Pasteur (1822-1895) và Robert Koch (1843-1910) miêu tả vai trò của vi khuẩn là các thể mang và gây ra bệnh hay tác nhân gây bệnh.
Ban đầu vi khuẩn hay vi trùng (microbe) được coi là các loại nấm có kích thước hiển vi (gọi là schizomycetes), ngoại trừ các loại vi khuẩn lam (cyanobacteria) quang hợp, được coi là một nhóm tảo (gọi là cyanophyta hay tảo lam). Phải đến khi có những nghiên cứu về cấu trúc tế bào thì vi khuẩn mới được nhìn nhận là một nhóm riêng khác với các sinh vật khác. Vào năm 1956 Hebert Copeland phân chúng vào một giới (kingdom) riêng là Mychota, sau đó được đổi tên thành Sinh vật khởi sinh (Monera), Sinh vật nhân sơ (Prokaryota), hay Vi khuẩn (Bacteria). Trong thập niên 1960, khái niệm này được xem xét lại và vi khuẩn (bây giờ gồm cả cyanbacteria) được xem như là một trong hai nhóm chính của sinh giới, cùng với sinh vật nhân chuẩn. Sinh vật nhân chuẩn được đa số cho là đã tiến hóa từ vi khuẩn, và sau đó cho rằng từ một nhóm vi khuẩn hợp lại.
Sự ra đời của phân loại học phân tử đã làm lung lay quan điểm này. Năm 1977, Carl Woese chia sinh vật nhân sơ thành 2 nhóm dựa trên trình tự 16S rRNA, gọi là vực Vi khuẩn chính thức (Eubacteria) và Vi khuẩn cổ Archaebacteria. Ông lý luận rằng hai nhóm này, cùng với sinh vật nhân chuẩn, tiến hóa độc lập với nhau và vào năm 1990 nhấn mạnh thêm quan điểm này bằng cách đưa ra hệ phân loại 3 vực (three-domain system), bao gồm Vi khuẩn (Bacteria), Vi khuẩn cổ (Archaea) và Sinh vật nhân chuẩn (Eucarya). Quan điểm này được chấp nhận rộng rãi giữa các nhà sinh học phân tử nhưng cũng bị chỉ trích bởi một số khác, cho rằng ông đã quan trọng hóa vài khác biệt di truyền và rằng cả vi khuẩn cổ và sinh vật nhân chuẩn có lẽ đều phát triển từ vi khuẩn chính thức.
Đặc điểm sinh sản
Vi khuẩn chỉ sinh sản vô tính (asexual reproduction), không sinh sản hữu tính (có tái tổ hợp di truyền). Cụ thể hơn, chúng sinh sản bằng cách chia đôi (binary fission), hay trực phân. Trong quá trình này, một tế bào mẹ được phân thành 2 tế bào con bằng cách tạo vách ngăn đôi tế bào mẹ.
Tuy nhiên, mặc dù không có sinh sản hữu tính, những biến đổi di truyền (hay đột biến) vẫn xảy ra trong từng tế bào vi khuẩn thông qua các hoạt động tái tổ hợp di truyền. Do đó, tương tự như ở các sinh vật bậc cao, kết quả cuối cùng là vi khuẩn cũng có được một tổ hợp các tính trạng từ hai tế bào mẹ. Có ba kiểu tái tổ hợp di truyền đã được phát hiện ở vi khuẩn:
biến nạp (transformation): chuyển DNA trần từ một tế bào vi khuẩn sang tế bào khác thông qua môi trường lỏng bên ngoài, hiện tượng này gồm cả vi khuẩn chết,
tải nạp (transduction): chuyển DNA của virus, vi khuẩn, hay cả virus lẫn vi khuẩn, từ một tế bào sang tế bào khác thông qua thể thực khuẩn (bacteriophage) và,
giao nạp (conjugation): chuyển DNA từ vi khuẩn này sang vi khuẩn khác thông qua cấu trúc protein gọi là pilus (lông giới tính).
Vi khuẩn, sau khi nhận được DNA từ một trong những cách trên, sẽ tiến hành phân chia và truyền bộ gene tái tổ hợp cho thế hệ sau. Nhiều vi khuẩn còn có plasmid chứa DNA nằm ngoài nhiễm sắc thể (extrachromosomal DNA). Dưới điều kiện thích hợp, vi khuẩn có thể tạo thành những khúm thấy được bằng mắt thường, chẳng hạn như bacterial mat.
Các vi khuẩn có rất nhiều kiểu trao đổi chất khác nhau.[16] Vi khuẩn dị dưỡng (heterotroph) phải dựa vào nguồn cacbon hữu cơ bên ngoài, và tất cả các vi khuẩn gây bệnh đều là các vi khuẩn dị dưỡng. Trong khi các vi khuẩn tự dưỡng (autotroph) có khả năng tổng hợp chất hữu cơ từ CO2 và nước. Các vi khuẩn tự dưỡng thu nhận năng lượng từ phản ứng ôxy hóa các hợp chất hóa học gọi là vi khuẩn hóa dưỡng (chemotroph), và những nhóm thu năng lượng từ ánh sáng, thông qua quá trình quang hợp, được gọi là vi khuẩn quang dưỡng (phototroph). Có nhiều cách khác để gọi hai nhóm theo thuật ngữ tiếng Anh, ví dụ như chemoautotroph và photosynthesis autotroph, v.v. Ngoài ra, các vi khuẩn còn được phân biệt nhờ vào nguồn chất khử mà chúng sử dụng. Những nhóm sử dụng hợp chất vô cơ (như nước, khí hiđrô, sulfua và ammonia) làm chất khử được gọi là vi khuẩn vô cơ dưỡng (lithotroph) và những nhóm cần hợp chất hữu cơ (như đường, axit hữu cơ) được gọi là vi khuẩn hữu cơ dưỡng (organotroph). Những kiểu trao đổi chất dựa vào nguồn năng lượng (quang dưỡng hay hóa dưỡng), nguồn chất khử (vô cơ dưỡng hay hữu cơ dưỡng) và nguồn cácbon (tự dưỡng hay dị dưỡng) có thể được kết hợp khác nhau trong từng tế bào, và nhiều loài có thể thường xuyên chuyển từ kiểu trao đổi chất này sang kiểu trao đổi chất khác.
Vi khuẩn quang vô cơ tự dưỡng bao gồm vi khuẩn lam (cyanobacteria), là một trong những loài cổ nhất được biết đến từ hóa thạch và có lẽ đã đóng một vai trò quan trọng trong việc tạo ra nguồn ôxy cho khí quyển Trái Đất. Chúng là những tiên phong trong việc sử dụng nước như là nguồn electron vô cơ (lithotrophic) và là sinh vật đầu tiên dùng bộ máy quang hợp để phân rã nước. Những vi khuẩn quang hợp khác dùng các nguồn electron khác nên không tạo ra ôxy. Những vi khuẩn quang dưỡng không tạo ôxy nằm trong bốn nhóm phân loại: vi khuẩn lục lưu huỳnh, vi khuẩn lục không dùng lưu huỳnh, vi khuẩn tía và heliobacteria.
Những chất dinh dưỡng cần thiết cho sự phát triển bình thường gồm nitơ, lưu hùynh, phôtpho, vitamin và các nguyên tố kim loại như natri, kali, canxi, ma-nhê, mangan, sắt, kẽm, côban, đồng, nikel... Vài loài cần thêm một số nguyên tố vết khác như selen, tungsten, vanađi hay bo.
Dựa vào phản ứng với ôxy, hầu hết các vi khuẩn có thể được xếp vào 3 nhóm: một số chỉ có thể mọc khi có sự hiện diện của ôxy được gọi là vi khuẩn hiếu khí (aerobe); một số khác chỉ có thể mọc khi không có ôxy được gọi là vi khuẩn kị khí (anaerobe); và một số lại có thể sống cả khi có hay không có sự hiện diện của ôxy thuộc nhóm vi khuẩn kị khí tùy ý (facultative anaerobe). Các vi khuẩn không sử dụng ôxy nhưng vẫn có thể mọc khi có sự hiện diện của ôxy gọi là vi khuẩn chịu oxy (aerotolerant). Những vi khuẩn có thể mọc tốt trong môi trường khắc nghiệt đối với con người được gọi là extremophile (vi khuẩn chịu cực hạn). Một số vi khuẩn sống trong suối nước nóng được gọi là vi khuẩn chịu nhiệt (thermophile); một số khác sống trong hồ nước rất mặn gọi là vi khuẩn chịu mặn (halophile); trong khi đó có loài lại sống trong môi trường acid hay kiềm gọi là vi khuẩn chịu axit (acidophile) hay vi khuẩn chịu kiềm (alkaliphile) và còn một số sống trong các băng hà trong dãy núi Alps gọi là vi khuẩn chịu hàn (psychrophile).
Di động
Vi khuẩn di động nhờ vào tiên mao (flagellum), trượt (bacterial gliding) hay thay đổi sức nổi (buoyancy). Một nhóm vi khuẩn đặc biệt, nhóm spirochaete, có các cấu trúc tương tự như tiên mao, gọi là sợi trục (axial filament), nằm giữa hai màng trong vùng chu chất. Chúng có một thể xoắn ốc đặc biệt quay tròn khi di chuyển.
Tiên mao của vi khuẩn được sắp xếp theo nhiều cách. Vi khuẩn có thể có một tiên mao ở mỗi cực của tế bào, hay có thể có một nhóm nhiều tiên mao ở một đầu. Peritrichous là nhóm vi khuẩn có tiên mao nằm rải rác khắp tế bào. Nhiều vi khuẩn (như E. coli) có hai kiểu di động khác nhau: di động tiến tới (bơi) và quay vòng. Di động quay vòng giúp chúng tái định hướng và là một nhân tố qua trọng tạo ra tính định hướng bất kì cho di động tiến tới.
Vi khuẩn di động bị thu hút hay đẩy ra bởi một số kích thích, hoạt động này được gọi là tính hướng động (taxes) chẳng hạn như hóa hướng động (chemotaxis), quang hướng động (phototaxis), cơ hướng động (mechanotaxis) và từ hướng động (magnetotaxis). Trong nhóm myxobacteria, các tế bào vi khuẩn có thể dính lại với nhau để tạo thành đám và có thể biệt hóa tạo thành thể quả.
Hình thái
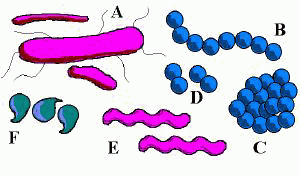
Vi khuẩn có nhiều hình dạng khác nhau:
A. Hình que - trực khuẩn (Bacillus)
B. Hình cầu (coccus) tạo thành chuỗi (strepto-) - liên cầu khuẩn (Streptococcus).
C. Hình cầu tạo đám (staphylo-) - tụ cầu khuẩn (Staphylococcus).
D. Hình tròn sóng đôi (diplo-) - song cầu khuẩn (Diplococcus).
E. Hình xoắn - xoắn khuẩn (Spirillum, Spirochete).
F. Hình dấu phẩy - phẩy khuẩn (Vibrio).
Vi khuẩn có nhiều hình dạng khác nhau. Các tế bào vi khuẩn chỉ bằng 1/10 tế bào của sinh vật nhân chuẩn và dài khoảng 0,5–5,0 micromet. Tuy nhiên, một vài loài như Thiomargarita namibiensis và Epulopiscium fishelsoni lại có kích chiều dài đến nửa mm và có thể nhìn thấy bằng mắt thường;[17] E. fishelsoni đạt 0,7 mm.[18] Những vi khuẩn nhỏ nhất là các thành viên thuộc chi Mycoplasma, chúng có kích thước chỉ 0,3 micromet, nhỏ bằng với virus lớn nhất.[19] Một số vi khuẩn thậm chí có thể nhỏ hơn, nhưng các vi khuẩn siêu nhỏ này chưa được nghiên cứu kỹ.[20]
Đa số có hình que, hình cầu, hay hình xoắn; những vi khuẩn có hình dạng như vậy được gọi theo thứ tự là trực khuẩn (bacillus), cầu khuẩn (coccus), và xoắn khuẩn (spirillum).[21] Một nhóm khác nữa là phẩy khuẩn (vibrio) có hình dấu phẩy. Một nhóm nhỏ hơn thậm chí có dạng hình tứ diện.[22] Các nghiên cứu gần đây phát hiện vi khuẩn ở sâu bên trong lớp vỏ Trái Đất, chúng có dạng sợi phân nhánh với mặt cắt ngang có hình sao. Diện tích bề mặt lớn so với tỉ số thể tích của dạng hình thái này có thể tạo cho các vi khuẩn này đặc điểm dễ thích nghi với môi trường nghèo dinh dưỡng.[23] Sự đa dạng về hình dạng được xác định thông qua thành tế bào và khung tế bào vi khuẩn, và đây là điều quan trong vì nó ảnh hưởng đến khả năng vi khuẩn có được chất dinh dưỡng, gắn vào các bề mặt, bơi trong chất lỏng và trốn kẻ săn mồi.[24][25] Hình dạng không còn được coi là một tiêu chuẩn định danh vi khuẩn, tuy nhiên có rất nhiều chi được đặt tên theo hình dạng (ví dụ như Bacillus, Streptococcus, Staphylococcus) và nó là một điểm quan trọng để nhận dạng các chi này.
Một công cụ quan trọng để nhận dạng khác là nhuộm Gram, đặt theo tên của Hans Christian Gram, người phát triển kĩ thuật này. Nhuộm Gram giúp phân vi khuẩn thành 2 nhóm, dựa vào thành phần cấu tạo của vách tế bào. Khi đầu tiên chính thức sắp xếp các vi khuẩn vào từng ngành, người ta dựa chủ yếu vào phản ứng này:
Gracilicutes - vi khuẩn có màng tế bào thứ cấp chứa lipid, nhuộm Gram âm tính (nói gọn là vi khuẩn Gram âm)
Firmicutes - vi khuẩn có một màng tế bào và vách pepticoglycan dày, nhuộm Gram cho kết quả dương tính (Gram dương)
Mollicutes - vi khuẩn không có màng thứ cấp hay vách, nhuộm Gram âm tính.
Các vi khuẩn cổ trước đây được xếp trong nhóm Mendosicutes. Như đã nói ở trên, ngành này không còn đại diện cho những nhóm có quan hệ tiến hóa nữa. Hầu hết vi khuẩn Gram dương được xếp vào ngành Firmicutes và Actinobacteria, là hai ngành có quan hệ gần. Tuy nhiên, ngành Firmicutes đã được định nghĩa lại và bao gồm cả mycoplasma (Mollicutes) và một số vi khuẩn Gram âm.
Vi khuẩn có thể có ích hoặc có hại cho môi trường, và động vật, bao gồm cả con người. Vai trò của vi khuẩn trong gây bệnh và truyền bệnh rất quan trọng. Một số là tác nhân gây bệnh (pathogen) và gây ra bệnh uốn ván (tetanus), sốt thương hàn (typhoid fever), giang mai (syphilis), tả (cholera), bệnh lây qua thực phẩm (foodborne illness) và lao (tuberculosis). Nhiễm khuẩn huyết (sepsis), là hội chứng nhiễm khuẩn toàn cơ thể gây sốc và giãn mạch, hay nhiễm khuẩn khu trú (localized infection), gây ra bởi các vi khuẩn như streptococcus, staphylococcus, hay nhiều loài Gram âm khác. Một số nhiễm khuẩn có thể lan rộng ra khắp cơ thể và trở thành toàn thân (systemic). Ở thực vật, vi khuẩn gây mụn lá (leaf spot), fireblight và héo cây. Các hình thức lây nhiễm gồm qua tiếp xúc, không khí, thực phẩm, nước và côn trùng. Kí chủ (host) bị nhiễm khuẩn có thể trị bằng thuốc kháng sinh, được chia làm hai nhóm là diệt khuẩn (bacteriocide) và kìm khuẩn (bacteriostasis), với liều lượng mà khi phân tán vào dịch cơ thể có thể tiêu diệt hoặc kìm hãm sự phát triển của vi khuẩn.
Các biện pháp khử khuẩn có thể được thực hiện để ngăn chặn sự lây lan của vi khuẩn, ví dụ như chùi da bằng cồn trước khi tiêm. Việc vô khuẩn các dụng cụ phẫu thuật và nha khoa được thực hiện để đảm bảo chúng "vô khuẩn" (sterile) hay không mang vi khuẩn gây bệnh, để ngăn chặn sự nhiễm khuẩn. Chất tẩy uế được dùng để diệt vi khuẩn hay các tác nhân gây bệnh để ngăn chặn sự nhiễm và nguy cơ nhiễm khuẩn.
Trong đất, các vi sinh vật sống trong nốt rễ (rhizosphere) biến nitơ thành ammoniac bằng các enzyme của chính mình. Một số khác lại dùng phân tử khí nitơ làm nguồn nitơ (đạm) cho mình, chuyển nitơ thành các hợp chất của nitơ, quá trình này gọi là quá trình cố định đạm. Nhiều vi khuẩn được tìm thấy sống cộng sinh trong cơ thể người hay các sinh vật khác. Ví dụ như sự hiện diện của các vi khuẩn cộng sinh trong ruột già giúp ngăn cản sự phát triển của các vi sinh vật có hại.
Vi khuẩn có khả năng phân giải các hợp chất hữu cơ một cách đáng kinh ngạc. Một số nhóm vi sinh "chuyên hóa" đóng một vai trò rất quan trọng trong việc hình thành các khoáng chất từ một số nhóm hợp chất hữu cơ. Ví dụ, sự phân giải cellulose, một trong những thành phần chiếm đa số trong mô thực vật, được thực hiện chủ yếu bởi các vi khuẩn hiếu khí thuộc chi Cytophaga. Khả năng này cũng được con người ứng dụng trong công nghiệp và trong cải thiện sinh học (bioremediation). Các vi khuẩn có khả năng phân hủy hydrocarbon trong dầu mỏ thường được dùng để làm sạch các vết dầu loang.
Vi khuẩn, cùng với nấm men và nấm mốc, được dùng để chế biến các thực phẩm lên men như phô-mai, dưa chua, nước tương, dưa cải bắp (sauerkraut), giấm, rượu, và yoghurt. Sử dụng công nghệ sinh học, các vi khuẩn có thể được "thiết kế" (bioengineer) để sản xuất thuốc trị bệnh như insulin, hay để cải thiện sinh học đối với các chất thải độc hại.
Các vấn đề khác
Về mặt tiến hóa học, vi khuẩn được cho là các vi sinh vật khá cổ, xuất hiện khoảng 3,7 tỉ năm trước.
Hai bào quan (organelle), ty thể (mitochondrion) và lục lạp (chloroplast), được đa số cho là bắt nguồn từ vi khuẩn nội cộng sinh (endosymbiotic). Xem thêm: thuyết nội cộng sinh.
Vi sinh vật phân bố khắp mọi nơi và phát triển nhanh chóng ở những nơi có đủ thức ăn, độ ẩm, và nhiệt độ tối ưu cho sự phân chia và lớn lên của chúng. Chúng có thể được mang đi bởi gió từ nơi này sang nơi khác. Cơ thể người là nơi cư trú của hằng tỷ vi sinh vật; chúng ở trên da, đường ruột, trong mũi, miệng và những nơi hở khác của cơ thể. Chúng có trong không khí mà ta thở, nước ta uống và thức ăn ta ăn.
Giới Khởi sinh (Monera) là một giới đã lỗi thời trong hệ thống năm giới của phân loại sinh học. Nó là những sinh vật nhỏ bé có kích thước hiển vi (từ 1-3μm) cấu tạo bởi các tế bào nhân sơ, là những sinh vật cổ sơ nhất xuất hiện khoảng 3,5 tỷ năm trước đây. Chúng sống khắp nơi, trong đất, nước, không khí; phương thức dinh dưỡng rất đa dạng: hoá tự dưỡng, hoá dị dưỡng, quang tự dưỡng và quang dị dưỡng. Nhiều vi khuẩn sống kí sinh trong các cơ thể khác. Vi khuẩn có chứa nhiều sắc tố quang hợp trong đó có diệp lục như vi khuẩn lam có khả năng tự dưỡng quang hợp như thực vật.
Giới Khởi sinh bao gồm phần lớn các sinh vật với cấu trúc tế bào nhân sơ. Vì lý do này nên giới Monera đôi khi cũng được gọi là Prokaryota hay Prokaryotae. Trước khi có sự tạo ra giới này thì nhóm sinh vật trong giới đã được coi như là thuộc về hai ngành tách rời của thực vật: Schizomycetes (vi khuẩn) đã được coi là nấm, và Cyanophyta được coi là tảo lục-lam. Nhóm cuối cùng này hiện nay được coi là một nhóm trong vi khuẩn, thông thường gọi là vi khuẩn lam và hiện tại đã biết là không có quan hệ họ hàng gần với thực vật, nấm hay động vật.
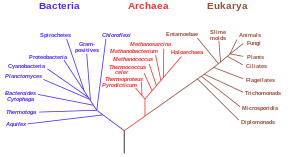
Các phân tích chuỗi gen ADN và ARN gần đây đã chứng minh rằng có hai nhóm chính của sinh vật nhân sơ là vi khuẩn (Bacteria) và vi khuẩn cổ (Archaea), chúng dường như không có mối quan hệ gần gũi hơn với nhau khi so với mối quan hệ của từng nhóm đối với sinh vật nhân chuẩn (Eukaryota/Eukarya). Vì thế, Monera kể từ đó đã bị chia ra thành Archaea và Bacteria, tạo thành hệ thống sáu giới và hệ thống ba vực gần đây. Tất cả các sơ đồ mới đều loại bỏ Monera và hiện nay coi Bacteria, Archaea, Eukarya như là ba vực (hay giới) tách rời.
Lịch sử
Theo truyền thống, sinh vật được phân loại như là động vật, thực vật, khoáng vật như trong Systema Naturae. Sau khi phát hiện ra kính hiển vi, các cố gắng được thực hiện nhằm xếp đặt vi sinh vật vào hoặc là giới thực vật hoặc là giới động vật. Năm 1866, Ernst Haeckel đề xuất hệ thống ba giới với sự bổ sung Protista như là giới mới và chứa phần lớn các vi sinh vật.[1] Sau đó, Haeckel đề xuất giới thứ tư mà ông gọi là Monera (từ tiếng Hy Lạp: μονήρης, moneres, nghĩa là đơn giản). Một trong tám ngành chính trong Protista của ông được gọi là Moneres. Tiểu thể loại Moneres của Haeckel bao gồm các nhóm vi khuẩn đã biết như Vibrio. Giới Protista của Haeckel cũng bao gồm các sinh vật với nhân chuẩn mà hiện nay người ta gọi là Protista (sinh vật nguyên sinh). Sau này người ta thấy rằng giới Protista của Haeckel là quá đa dạng để có thể coi một cách nghiêm túc như là một giới thật sự.
Năm 1969, Robert Whittaker công bố hệ thống năm giới do ông đề xuất để phân loại các sinh vật.[2] Hệ thống của Whittaker đặt phần lớn các sinh vật đơn bào vào trong hoặc là Monera với nhân sơ hay Protista với nhân chuẩn. Ba giới còn lại trong hệ thống của ông là Fungi, Animalia, và Plantae với nhân chuẩn
Phân chia Monera
Cây phát sinh loài, dựa trên các dữ liệu chuỗi rARN, chỉ ra sự tách rời của vi khuẩn, vi khuẩn cổ và sinh vật nhân chuẩn.
Dựa trên các nghiên cứu phát sinh loài ở mức phân tử, Carl Woese đã đề xuất rằng các sinh vật nhân sơ (Monera) được chia thành 2 nhóm tách rời là Bacteria và Archaea. Trong phát sinh loài do Carl Woese đề xuất năm 1990[3], cả ba giới này đều có nguồn gốc từ một tổ tiên chung toàn thế giới và điều này là phát sinh loài thể loại được chấp nhận phổ biến nhất ngày nay. Tuy nhiên, diễn giải hiện đại nhất cho ba giới này là "Universal and Eukaryote Phylogenetic Tree" (Cây phát sinh loài chung và sinh vật nhân chuẩn) dựa trên rADN 16s, như được thể hiện trong Tree of Life Web Project (Dự án sơ đồ của các sinh vật sống).[4]
Vi khuẩn và vi khuẩn cổ
Điểm khác nhau đáng chú ý nhất giữa vi khuẩn thật sự (Eubacteria) và vi khuẩn cổ (Archaebacteria) là các môi trường mà chúng có thể sinh sống. Vi khuẩn thật sự chiếm phần lớn trong vi khuẩn mà con người có thể tiếp xúc. Những vi khuẩn sống trong hay xung quanh con người, như Escherichia coli và các loài từ chi Salmonella, là vi khuẩn thật sự. Vi khuẩn cổ sinh sống trong các môi trường khắc nghiệt hơn, chẳng hạn trong các suối nước nóng có độ axít cao hay tại các độ sâu hàng dặm dưới lớp băng Bắc cực. Các nhóm này sau đó đã được đổi tên thành Bacteria và Archaea.
Động vật là một nhóm sinh vật đa bào, nhân chuẩn, được phân loại là giới Động vật (Animalia, đồng nghĩa:Metazoa) trong hệ thống phân loại 5 giới. Cơ thể của chúng lớn lên khi phát triển. Hầu hết động vật có khả năng di chuyển một cách tự nhiên và độc lập.
Hầu hết các ngành động vật được biết đến nhiều nhất đã xuất hiện hóa thạch vào thời kỳ Bùng nổ kỷ Cambri, khoảng 542 triệu năm trước. Động vật được chia thành nhiều nhóm nhỏ, một vài trong số đó là động vật có xương sống (chim, động vật có vú, lưỡng cư, bò sát, cá); động vật thân mềm (trai, hàu, bạch tuộc, mực, và ốc sên); động vật Chân khớp (cuốn chiếu, rết, côn trùng, nhện, bọ cạp, tôm hùm, tôm); giun đốt (giun đất, đỉa); bọt biển và sứa.
Theo chiều kim đồng hồ từ phía trên mé trái: Mực ống châu Âu (Loligo vulgaris - động vật thân mềm), Chrysaora quinquecirrha - sứa lông châm, Arthropoda - động vật chân khớp, Eunereis longissima -giun đốt, hổ (Panthera tigris - động vật có dây sống).
Từ nguyên (tên gọi)
Từ "animal" xuất phát từ tiếng Latin animalis, có nghĩa là "có thở".[1] Trong sử dụng ngôn ngữ giao tiếp hàng ngày, từ "động vật" thường bị sử dụng sai - từ "động vật" đó dùng để chỉ tất cả các thành viên của giới Animalia trừ con người. Theo nghĩa sinh học, "động vật" dùng để chỉ tất cả các thành viên của giới Animalia, bao gồm cả con người.[2]
Đặc điểm
Động vật có vài đặc điểm riêng tách chúng ra khỏi các sinh vật sống khác. Động vật là sinh vật nhân chuẩn và đa bào,[3] tách chúng khỏi vi khuẩn và hầu hết sinh vật đơn bào. Động vật sống dị dưỡng,[4] tiêu hóa thức ăn trong cơ thể, tách chúng ra khỏi thực vật và tảo.[5] Chúng cũng khác biệt với thực vật ở chỗ thiếu thành tế bào cứng.[6] Tất cả động vật có thể di chuyển,[7] ít nhất là trong một giai đoạn sống. Ở hầu hết động vật, phôi trải qua giai đoạn phôi nang (blastula),[8] một gai đoạn riêng biệt đặc trưng ở động vật.
Cấu trúc
Trừ vài ngoại lệ, như là bọt biển (ngành Porifera) và Placozoa, động vật có cơ thể được chia thành các mô. Chúng bao gồm cơ bắp, dùng để thực hiện và kiểm soát vận động, các mô thần kinh, dùng để gửi và sử lí tính hiệu. Thông thường, cơ thể có một hệ tiêu hóa, với một miệng (như thủy tức) hay cả miệng và hậu môn (như cá).[9] Tất cả động vật có tế bào nhân chuẩn.
Sinh sản và phát triển
Một tế bào phổi sa giông được nhuộm huỳnh quang trải qua giai đoạn đầu pha sau phân bào
Gần như tất cả động vật trải qua một số hình thức sinh sản hữu tính.[10] Chúng có những tế bào sinh sản nhỏ, di chuyển được như tinh trùng hay lớn hơn, không di chuyển được như trứng.[11] Tình trùng và trứng sẽ kết hợp để tạo thành hợp tử, hợp tử phát triển để tạo thành cá thể mới.[12]
Nhiều loài động vật cũng có khả năng sinh sản vô tính.[13] Việc này có thể xảy ra thông qua trinh sản, trứng được tạo ra mà không cần giao phối, phân chồi, hay phân mảnh.[14]
Hợp tử ban đầu phát triển thành một khối tế bào hình cầu rỗng, được gọi là phôi nang,[15] sau sẽ được sắp xếp lại. Ở bọt biển, ấu trùng phôi nang bơi đến một vị trí mới và phát triển thành một con bọt biển mới.[16] Trong các nhóm khác, phôi nang trải qua những sắp xếp phức tạp hơn.[17]
Nguồn gốc và hóa thạch
Động vật thường được coi là tiến hóa từ một loại trùng roi có tế bào nhân chuẩn.[18] Họ hàng gần gũi nhất được biết đến của chúng là Choanoflagellatea. Nghiên cứu phân tử đặt động vật trong một siêu nhóm được gọi là opisthokonta (sinh vật lông roi sau), cùng với choanoflagellate, nấm và một số sinh vật nguyên sinh ký sinh nhỏ.[19] Tên này đến từ vị trí của roi trong tế bào có thể chuyển động, như tinh trùng ở hầu hết động vật, trong khi các sinh vật nhân chuẩn khác có lông roi trước.[20]
Những hóa thạch đầu tiên được cho có thể là động vật xuất hiện ở thành hệ Trezona, tây Central Flinders, Nam Úc.[21] Những hóa thạch này được xem là loài bọt biển đầu tiên. Chúng được tìm thấy trong lớp đá 665 triệu năm tuổi.[21]
Hóa thạch tiếp theo có thể là động vật cổ nhất được tìm thấy vào thời kỳ Tiền Cambri, khoảng 610 triệu năm trước.[22] Hóa thạch này khó mà liên quan đến các hóa thạch sau nó. Tuy nhiên, hóa thạch này có thể đại diện cho động vật tiền thân của động vật ngày nay, nhưng chúng cũng có thể là một nhóm tách biệt hoặc thậm chí không phải động vật thực sự.[23]
Lịch sử phân loại
Carolus Linnaeus, được biết đến như là cha đẻ của phân loại hiện đại
Aristotle chia sinh vật sống ra làm động vật và thực vật, Carolus Linnaeus (Carl von Linné) cũng làm theo cách này trong lần phân loại thứ bậc đầu tiên.[24] Kể từ đó các nhà sinh học đã bắt đầu nhấn mạnh mối quan hệ tiến hóa, ví dụ sinh vật đơn bào ban đầu được xem là động vật bởi khả năng di chuyển của chúng, nhưng nay được tách riêng.
Trong sơ đồ ban đầu của Linnaeus, động vật là một trong ba giới, phân chia thành các lớp Vermes, Insecta, Pisces, Amphibia, Aves, và Mammalia. Kể từ đó, bốn lớp cuối được gộp thành một ngành duy nhất, Chordata, trong khi hai lớp còn lại bị tách ra.
Virus (phiên âm là vi-rút), còn gọi là siêu vi, siêu vi khuẩn hay siêu vi trùng, là một tác nhân truyền nhiễm chỉ nhân lên được khi ở bên trong tế bào sống của một sinh vật khác. Virus có thể xâm nhiễm vào tất cả các dạng sinh vật, từ động vật, thực vật cho tới vi khuẩn và vi khuẩn cổ.[1] Kể từ bài viết đầu tiên của D. I. Ivanovskiy năm 1892, mô tả về một dạng mầm bệnh không thuộc vi khuẩn mà lây nhiễm vào cây thuốc lá, và sự khám phá ra virus khảm thuốc lá của Martinus Beijerinck năm 1898,[2] cho đến nay có khoảng 5.000 loại virus đã được miêu tả chi tiết,[3] mặc dù vẫn còn có tới hàng triệu dạng virus khác nhau.[4] Virus được tìm thấy ở hầu hết mọi hệ sinh thái trên Trái Đất và là dạng có số lượng nhiều nhất trong tất cả các thực thể sinh học.[5][6] Khoa học nghiên cứu virus được biết với tên virus học (virology), một chuyên ngành phụ của vi sinh vật học.
Các phần tử (hay hạt) virus (được gọi là virion) được tạo thành từ hai hoặc ba bộ phận: i) phần vật chất di truyền được tạo nên từ ADN hoặc ARN, là những phân tử dài có mang thông tin di truyền; ii) một lớp vỏ protein - được gọi với tên capsid - có chức năng bảo vệ hệ gen; và trong một số trường hợp còn có iii) một lớp vỏ bọc bên ngoài làm từ lipid mà bao bọc bên ngoài lớp vỏ protein khi virus ở ngoài tế bào. Hình dạng của virus có sự khác nhau, từ dạng xoắn ốc hay khối hai mươi mặt đều đơn giản cho tới những cấu trúc phức tạp hơn. Một virus có kích thước trung bình vào khoảng 1/100 kích cỡ trung bình của một vi khuẩn. Hầu hết virus đều quá nhỏ nên không thể quan sát trực tiếp dưới kính hiển vi quang học.
Nguồn gốc của virus trong lịch sử tiến hóa của sự sống không rõ ràng: một số có thể đã tiến hóa từ những plasmids – những đoạn ADN ngắn có khả năng di chuyển giữa các tế bào – trong khi số khác có thể đã tiến hóa từ vi khuẩn. Trong tiến hóa, virus là một phương tiện chuyển gen ngang quan trọng, góp phần gia tăng sự đa dạng di truyền.[7] Virus được công nhận là một dạng sống bởi một số nhà khoa học, do chúng có mang vật chất di truyền, có thể sinh sản và tiến hóa thông qua quá trình chọn lọc tự nhiên. Tuy nhiên mặt khác chúng lại thiếu những đặc tính thiết yếu (như cấu trúc tế bào) - những điều được công nhận rộng rãi là cần thiết để được coi như sinh vật sống. Bởi vì chỉ có một số chứ không tất cả các phẩm chất cần thiết, nên virus được mô tả như "những sinh vật ở bên lề của sự sống".[8]
Virus lây lan theo nhiều cách; virus thực vật thường được truyền từ cây này sang cây khác qua những loài côn trùng hút nhựa cây như rệp vừng; trong khi virus động vật lại có thể được truyền đi nhờ những côn trùng hút máu. Những sinh vật mang mầm bệnh như vậy được gọi là những vector. Virus cúm lan truyền thông qua ho và hắt hơi. Norovirus và rotavirus, nguyên nhân chính của bệnh viêm dạ dày-ruột siêu vi, lây lan qua đường phân-miệng và truyền từ người này sang người khác thông qua tiếp xúc, cũng như xâm nhập vào cơ thể qua thức ăn hay nước uống. HIV là một trong vài loại virus lây nhiễm thông qua quan hệ tình dục và tiếp xúc với máu bị nhiễm bệnh. Mỗi virus chỉ có thể xâm nhiễm vào một số dạng tế bào vật chủ nhất định, gọi là "biên độ vật chủ" (host range); biên độ này có thể rất hẹp hoặc rất rộng, tùy vào số lượng những sinh vật khác nhau mà virus có khả năng lây nhiễm.[9]
Sự xâm nhập của virus trong động vật đã kích hoạt một phản ứng miễn dịch nhằm loại bỏ virus xâm nhiễm. Những phản ứng miễn dịch cũng có thể được tạo ra bởi vắc-xin, giúp tạo ra miễn dịch thu được nhân tạo đối với một virus xâm nhiễm nhất định. Tuy nhiên, một số virus, bao gồm những loại gây ra AIDS và viêm gan siêu vi, lại có thể trốn tránh những phản ứng trên và gây ra sự nhiễm bệnh mãn tính. Đa phần các chất kháng sinh không có hiệu quả đối với virus, dù vậy cũng đã có những loại thuốc kháng virus được phát triển.
Từ nguyên học
Virus là một từ có xuất xứ từ tiếng Latin dùng để chỉ chất độc, được sử dụng lần đầu tại Anh vào năm 1392.[10] Tính từ Virulent, bắt nguồn từ tiếng Latin virulentus (độc hại), xuất hiện từ 1400.[11] Virus được ghi nhận với nghĩa "tác nhân gây bệnh truyền nhiễm" lần đầu vào năm 1728,[10] trước khi Dmitri Ivanovsky phát hiện ra virus vào năm 1892. Thuật ngữ virion bắt đầu có từ năm 1959,[12] được dùng để chỉ một phần tử virus, đơn lẻ, ổn định, có khả năng lây nhiễm mà được giải phóng ra từ tế bào và hoàn toàn có thể xâm nhập các tế bào khác cùng loại.[13]
Trong tiếng Việt, virus còn có tên gọi và cách viết khác là vi-rút, virut, siêu vi khuẩn hay siêu vi trùng.
Martinus Beijerinck trong phòng thí nghiệm vào năm 1921.
Vào những năm 1880, Louis Pasteur khi nghiên cứu về bệnh dại đã không thể tìm ra tác nhân gây ra bệnh này, và dự đoán về một mầm bệnh mà quá nhỏ để có thể phát hiện được dưới kính hiển vi.[14] Năm 1884, nhà vi sinh vật học người Pháp Charles Chamberland đã phát minh ra một bộ lọc (được biết tới ngày nay là bộ lọc Chamberland hay bộ lọc Chamberland-Pasteur) với các lỗ có kích thước còn nhỏ hơn cả vi khuẩn. Nhờ thế, ông có thể cho một dung dịch chứa vi khuẩn chảy qua bộ lọc và hoàn toàn loại bỏ chúng khỏi dung dịch.[15] Năm 1892, nhà sinh vật học người Nga, Dmitriy Iosifovich Ivanovskiy đã sử dụng bộ lọc này để nghiên cứu về thứ mà hiện nay được biết với tên virus khảm thuốc lá. Thí nghiệm của ông cho thấy chiết xuất từ lá cây thuốc lá nhiễm bệnh được nghiền nát vẫn có thể lây nhiễm sau khi lọc. D. I. Ivanovskiy đề xuất rằng sự nhiễm bệnh có thể là do một độc tố từ vi khuẩn gây ra, nhưng đã không theo đuổi ý tưởng đó.[16] Lý do là vì vào thời điểm đó, người ta đã nghĩ rằng tất cả các tác nhân truyền nhiễm đều có thể bị các bộ lọc giữ lại và chỉ có thể phát triển trên một môi trường dinh dưỡng – đây là một phần của thuyết mầm bệnh.[2] Năm 1898, nhà vi sinh người Hà Lan Martinus Beijerinck đã lặp lại thí nghiệm và tin rằng dung dịch đã lọc vẫn còn chứa một dạng tác nhân truyền nhiễm mới.[17] Ông nhận thấy rằng tác nhân này chỉ có thể nhân lên trong tế bào mà đang phân chia, nhưng thí nghiệm của ông không chỉ ra là nó được làm từ các hạt; ông gọi tác nhân này là một contagium vivum fluidum (mầm sống có thể hòa tan) và sử dụng lại từ virus để gọi nó.[16] Beijerinck giữ quan điểm rằng virus có bản chất là chất lỏng, tuy nhiên về sau thuyết này đã bị bác bỏ bởi Wendell Stanley, người chứng minh được chúng có dạng hạt.[16] Trong cùng năm đó, Friedrich Loeffler và Frosch đã cho chảy lần đầu tiên dung dịch chứa virus động vật – tác nhân gây bệnh lở mồm long móng (aphthovirus) – qua một bộ lọc tương tự.[18]
Vào đầu của thế kỷ 20, Frederick Twort - một nhà vi khuẩn học người Anh - đã khám phá ra một nhóm những virus mà xâm nhiễm vào vi khuẩn, mà nay gọi là thực khuẩn thể (bacteriophage hay phage),[19] và nhà vi sinh học người Canada gốc Pháp Félix d'Herelle đã miêu tả về virus rằng: khi thêm chúng vào vi khuẩn trên thạch agar, sẽ tạo ra những vùng vi khuẩn bị chết. Ông đã pha loãng chính xác một dịch huyền phù những virus trên và khám phá ra rằng những dịch pha loãng cao nhất (mật độ virus thấp nhất), thay vì giết chết toàn bộ vi khuẩn, đã tạo những vùng riêng biệt gồm những cá thể bị chết. Tính toán diện tích những vùng này và nhân với hệ số pha loãng cho phép ông tính được số lượng virus trong dịch huyền phù gốc.[20] Phage đã được báo trước sẽ là một giải pháp điều trị tiềm năng cho những bệnh như thương hàn và tả, nhưng triển vọng của chúng đã bị lãng quên cùng với sự phát triển của penicillin. Nghiên cứu về phage đã cung cấp cái nhìn sâu hơn về sự bất hoạt và kích hoạt gen, và một cơ chế hữu hiệu cho việc đưa những gen bên ngoài vào bên trong vi khuẩn.
Cho đến cuối thế kỷ 19, virus được định nghĩa dựa trên sự lây nhiễm của chúng, khả năng chống lọc, và việc chúng đòi hỏi phải có một vật chủ. Virus đã từng chỉ được nuôi trong thực vật và động vật. Năm 1906, Ross Granville Harrison phát minh ra một phương pháp để nuôi dưỡng mô trong bạch huyết, và sau đó năm 1913, E. Steinhardt, C. Israeli, và R. A. Lambert đã sử dụng phương pháp này để phát triển virus vaccinia trong những mảnh vụn của mô giác mạc chuột lang nhà.[21] Năm 1928, H. B. Maitland và M. C. Maitland đã nuôi virus vaccinia trong những thể huyền phù của thận gà băm nhỏ. Phương pháp của họ đã không được áp dụng rộng rãi cho tới thập niên 1950, khi poliovirus được nuôi ở quy mô lớn phục vụ việc sản xuất vaccine.[22]
Một bước đột phá khác đến vào năm 1931, khi nhà bệnh học Hoa Kỳ Ernest William Goodpasture đã nuôi dưỡng virus cúm và vài loại virus khác trong trứng gà đã thụ tinh.[23] Năm 1949, John Franklin Enders, Thomas Weller, và Frederick Robbins cũng nuôi cấy virus bại liệt trong tế bào phôi người, virus đầu tiên được nuôi mà không sử dụng mô thể rắn của động vật hay trứng. Công trình này cho phép Jonas Salk tạo ra một vắc-xin bại liệt hiệu quả.[24]
Những hình ảnh đầu tiên của virus thu nhận được là nhờ sự phát minh ra kính hiển vi điện tử năm 1931 của hai kĩ sư người Đức Ernst Ruska và Max Knoll.[25] Năm 1935, Wendell Meredith Stanley - một nhà sinh hóa và virus học người Mỹ - đã nghiên cứu virus khảm thuốc lá và nhận thấy chúng được tạo thành phần lớn từ protein.[26] Một thời gian ngắn sau, virus này đã được phân tách thành các phần protein và ARN riêng biệt.[27] Virus khảm thuốc lá là dạng virus đầu tiên được tinh thể hóa và cấu trúc của nó do đó đã được làm sáng tỏ chi tiết. Những hình ảnh nhiễu xạ tia X đầu tiên của virus kết tinh đã được Bernal và Fankuchen thu được vào 1941. Dựa trên những tấm hình này, Rosalind Franklin đã khám phá ra cấu trúc ADN hoàn thiện của loại virus này vào năm 1955.[28] Cùng trong năm đó, Heinz Fraenkel-Conrat và Robley Williams chứng minh được là chiết xuất ARN và vỏ protein của virus khảm thuốc lá có thể tự lắp ráp lại để tạo thành những virus có chức năng, cho thấy cơ chế đơn giản này có thể là cách virus sinh ra trong tế bào vật chủ.[29]
Nửa sau của thế kỷ 20 đánh dấu một kỉ nguyên vàng cho sự khám phá virus với hầu hết trong số hơn 2.000 loài virus động vật, thực vật và vi khuẩn đã được phát hiện trong những năm này.[30] Năm 1957, virus arteri ở ngựa và virus gây bệnh tiêu chảy ở bò (một loại pestivirus) đã đươc phát hiện. Năm 1963, virus viêm gan siêu vi B cũng được Baruch Blumberg khám phá,[31] và năm 1965, Howard Temin đã mô tả loại retrovirus đầu tiên. Sau đó, enzym phiên mã ngược (Reverse transcriptase), loại enzym quan trọng mà retrovirus sử dụng để phiên mã ARN của chúng thành ADN, được miêu tả lần đầu vào năm 1970, một cách độc lập bởi Howard Martin Temin và David Baltimore.[32] Năm 1983, nhóm nghiên cứu của Luc Montagnier tại Viện Pasteur ở Pháp, đã lần đầu tiên phân lập một loại retrovirus được biết với tên gọi ngày nay là virus suy giảm miễn dịch ở người (HIV).[33]
Nguồn gốc
Virus được tìm thấy ở bất cứ nơi nào có sự sống và có thể đã tồn tại kể từ khi tế bào sống đầu tiên được tiến hóa thành.[34] Nguồn gốc của virus không rõ ràng bởi chúng không tạo hóa thạch, vì vậy các kĩ thuật phân tử đã được sử dụng để so sánh ARN hay ADN của virus và là một công cụ hiệu quả để nghiên cứu xem chúng phát sinh ra sao.[35] Nhìn chung, có ba giả thuyết chính để giải thích nguồn gốc của virus:[36][37]
Giả thuyết thoái lui (regressive hypothesis)
Giả thuyết này cho rằng virus có thể đã từng là những tế bào nhỏ mà ký sinh bên trong những tế bào lớn hơn. Trải qua thời gian, những gen không cần thiết cho sự ký sinh này mất đi. Những vi khuẩn như Rickettsia và Chlamydia cũng sống trong tế bào và giống như virus, chỉ có thể sinh sản khi ở bên trong tế bào vật chủ. Những vi khuẩn này đã hỗ trợ cho giả thuyết thoái lui, do sự phụ thuộc ký sinh của chúng có thể đã làm mất đi những gen cho phép chúng tồn tại bên ngoài tế bào. Giả thuyết này còn được gọi là "giả thuyết thoái hóa" (degeneracy hypothesis),[38][39] hoặc "giả thuyết suy giảm" (reduction hypothesis).[40] Giả thuyết nguồn gốc từ tế bào (Cellular origin hypothesis)
Theo giả thuyết này, một số virus có thể đã tiến hóa từ những mảnh ADN hay ARN mà "thoát ra" (escape) từ hệ gen của những sinh vật lớn hơn. ADN thoát ra có thể là từ những plasmid (những đoạn ADN trần mà có thể di chuyển giữa những tế bào) hoặc từ những transposon (những phân tử ADN mà nhân lên và di chuyển quanh những vị trí khác nhau bên trong bộ gen của tế bào).[41] Từng được gọi là những "gen nhảy", transposon là những ví dụ của các yếu tố di truyền di động và có thể là nguồn gốc của một số virus. Chúng được phát hiện ở ngô bởi Barbara McClintock vào năm 1950.[42] Đôi khi giả thuyết này còn được gọi là "giả thuyết lang thang" (vagrancy hypothesis),[38][43] hoặc "giả thuyết trốn thoát" (escape hypothesis).[40] Giả thuyết đồng tiến hóa (coevolution hypothesis)
còn được gọi là "giả thuyết virus-đầu tiên" (virus-first hypothesis),[40] đề xuất rằng virus có thể đã tiến hóa từ những phân tử protein và axít nucleic phức tạp cùng một thời điểm khi tế bào xuất hiện lần đầu trên Trái Đất, và đã không hề bị phụ thuộc vào tế bào trong hàng tỷ năm. Viroid là những phân tử ARN mà không được phân loại là virus bởi chúng thiếu đi lớp vỏ protein. Tuy nhiên, chúng có những đặc tính tương đồng với vài loại virus và thường được gọi là những "tác nhân dưới virus" (subviral agent).[44] Viroid là những mầm bệnh quan trọng ở thực vật.[45] Chúng không mã hóa protein nhưng lại tương tác với tế bào chủ và sử dụng bộ máy tế bào vật chủ cho sự nhân lên của chúng.[46] Virus viêm gan siêu vi D trên người cũng có bộ gen ARN tương đồng với viroid nhưng có lớp vỏ protein xuất xứ từ virus viêm gan B và không có khả năng tạo ra lớp vỏ của riêng chúng. Vì vậy, chúng là một loại virus khiếm khuyết và không thể tự nhân lên mà không có virus viêm gan B giúp đỡ.[47] Tương tự như vậy, sputnik virophage cũng bị lệ thuộc vào mimivirus, loại virus lây nhiễm trên Acanthamoeba castellanii.[48] Những virus mà phụ thuộc vào sự hiện diện của những loài virus khác trong tế bào vật chủ được gọi là những vệ tinh và có thể là đại diện cho một bước trung gian trong quá trình tiến hóa giữa viroid và virus.[49][50]
Trong quá khứ, tất cả những giả thuyết trên đều gặp phải vấn đề: giả thuyết thoái lui không giải thích được tại sao kể cả những ký sinh nội bào nhỏ nhất cũng không giống với virus ở bất kỳ góc độ nào. Giả thuyết trốn thoát không lý giải được về lớp vỏ capsid phức hợp và các cấu trúc khác của phần tử virus. Giả thuyết virus-đầu tiên thì trái với định nghĩa của virus là chúng đòi hỏi phải có tế bào chủ.[40] Cho đến hiện nay, virus được công nhận là rất cổ xưa và có nguồn gốc mà bắt đầu từ trước cả sự rẽ nhánh của sự sống vào ba vực.[51] Phát hiện này đã khiến những nhà virus học hiện đại phải xem xét và đánh giá lại cả ba giả thuyết cổ điển trên.[51]
Bằng chứng về một thế giới của những tế bào ARN tổ tiên[52] và những phân tích máy tính về trình tự ADN của virus và vật chủ đã đem lại sự hiểu biết tốt hơn về mối quan hệ tiến hóa giữa những loại virus khác nhau, và có thể giúp xác định tổ tiên của những virus hiện đại. Cho đến nay, những phân tích này vẫn chưa chứng tỏ được giả thuyết nào ở trên là đúng.[52] Tuy nhiên, có vẻ như ít có khả năng rằng tất cả các virus hiện đang được biết có cùng một tổ tiên chung, và virus có lẽ đã phát sinh nhiều lần trong quá khứ bởi một hay nhiều cơ chế.[53]
Prion là những phân tử protein có khả năng lây nhiễm mà không phải ADN hay ARN.[54] Chúng có thể gây các bệnh như bệnh ngứa điên ở cừu (scrapie), bệnh viêm não thể bọt biển ở bò (bệnh bò điên) và bệnh suy mòn mãn tính ở hươu; ở con người, những bệnh gây ra bởi prion bao gồm bệnh Kuru, bệnh Creutzfeldt–Jakob, và hội chứng Gerstmann–Sträussler–Scheinker.[55] Prion có khả năng nhân lên do một số protein có thể tồn tại ở hai hình dạng khác nhau, và prion thay đổi hình dạng thông thường của protein vật chủ thành dạng prion. Điều này sẽ phát động một phản ứng chuỗi khi mỗi protein prion biến đổi nhiều protein vật chủ thành nhiều prion hơn, và những prion mới này lại tiếp tục biến đổi càng lúc càng nhiều protein thành prion hơn nữa; và tất cả những bệnh liên quan tới prion được biết đến đều dẫn đến tử vong. Mặc dù những prion về cơ bản khá khác biệt với virus và viroid, thì sự khám phá ra chúng đã làm gia tăng niềm tin vào học thuyết rằng virus có thể đã tiến hóa từ những phân tử tự sao chép.[56]
Martinus Beijerinck trong phòng thí nghiệm vào năm 1921.
Vào những năm 1880, Louis Pasteur khi nghiên cứu về bệnh dại đã không thể tìm ra tác nhân gây ra bệnh này, và dự đoán về một mầm bệnh mà quá nhỏ để có thể phát hiện được dưới kính hiển vi.[14] Năm 1884, nhà vi sinh vật học người Pháp Charles Chamberland đã phát minh ra một bộ lọc (được biết tới ngày nay là bộ lọc Chamberland hay bộ lọc Chamberland-Pasteur) với các lỗ có kích thước còn nhỏ hơn cả vi khuẩn. Nhờ thế, ông có thể cho một dung dịch chứa vi khuẩn chảy qua bộ lọc và hoàn toàn loại bỏ chúng khỏi dung dịch.[15] Năm 1892, nhà sinh vật học người Nga, Dmitriy Iosifovich Ivanovskiy đã sử dụng bộ lọc này để nghiên cứu về thứ mà hiện nay được biết với tên virus khảm thuốc lá. Thí nghiệm của ông cho thấy chiết xuất từ lá cây thuốc lá nhiễm bệnh được nghiền nát vẫn có thể lây nhiễm sau khi lọc. D. I. Ivanovskiy đề xuất rằng sự nhiễm bệnh có thể là do một độc tố từ vi khuẩn gây ra, nhưng đã không theo đuổi ý tưởng đó.[16] Lý do là vì vào thời điểm đó, người ta đã nghĩ rằng tất cả các tác nhân truyền nhiễm đều có thể bị các bộ lọc giữ lại và chỉ có thể phát triển trên một môi trường dinh dưỡng – đây là một phần của thuyết mầm bệnh.[2] Năm 1898, nhà vi sinh người Hà Lan Martinus Beijerinck đã lặp lại thí nghiệm và tin rằng dung dịch đã lọc vẫn còn chứa một dạng tác nhân truyền nhiễm mới.[17] Ông nhận thấy rằng tác nhân này chỉ có thể nhân lên trong tế bào mà đang phân chia, nhưng thí nghiệm của ông không chỉ ra là nó được làm từ các hạt; ông gọi tác nhân này là một contagium vivum fluidum (mầm sống có thể hòa tan) và sử dụng lại từ virus để gọi nó.[16] Beijerinck giữ quan điểm rằng virus có bản chất là chất lỏng, tuy nhiên về sau thuyết này đã bị bác bỏ bởi Wendell Stanley, người chứng minh được chúng có dạng hạt.[16] Trong cùng năm đó, Friedrich Loeffler và Frosch đã cho chảy lần đầu tiên dung dịch chứa virus động vật – tác nhân gây bệnh lở mồm long móng (aphthovirus) – qua một bộ lọc tương tự.[18]
Vào đầu của thế kỷ 20, Frederick Twort - một nhà vi khuẩn học người Anh - đã khám phá ra một nhóm những virus mà xâm nhiễm vào vi khuẩn, mà nay gọi là thực khuẩn thể (bacteriophage hay phage),[19] và nhà vi sinh học người Canada gốc Pháp Félix d'Herelle đã miêu tả về virus rằng: khi thêm chúng vào vi khuẩn trên thạch agar, sẽ tạo ra những vùng vi khuẩn bị chết. Ông đã pha loãng chính xác một dịch huyền phù những virus trên và khám phá ra rằng những dịch pha loãng cao nhất (mật độ virus thấp nhất), thay vì giết chết toàn bộ vi khuẩn, đã tạo những vùng riêng biệt gồm những cá thể bị chết. Tính toán diện tích những vùng này và nhân với hệ số pha loãng cho phép ông tính được số lượng virus trong dịch huyền phù gốc.[20] Phage đã được báo trước sẽ là một giải pháp điều trị tiềm năng cho những bệnh như thương hàn và tả, nhưng triển vọng của chúng đã bị lãng quên cùng với sự phát triển của penicillin. Nghiên cứu về phage đã cung cấp cái nhìn sâu hơn về sự bất hoạt và kích hoạt gen, và một cơ chế hữu hiệu cho việc đưa những gen bên ngoài vào bên trong vi khuẩn.
Cho đến cuối thế kỷ 19, virus được định nghĩa dựa trên sự lây nhiễm của chúng, khả năng chống lọc, và việc chúng đòi hỏi phải có một vật chủ. Virus đã từng chỉ được nuôi trong thực vật và động vật. Năm 1906, Ross Granville Harrison phát minh ra một phương pháp để nuôi dưỡng mô trong bạch huyết, và sau đó năm 1913, E. Steinhardt, C. Israeli, và R. A. Lambert đã sử dụng phương pháp này để phát triển virus vaccinia trong những mảnh vụn của mô giác mạc chuột lang nhà.[21] Năm 1928, H. B. Maitland và M. C. Maitland đã nuôi virus vaccinia trong những thể huyền phù của thận gà băm nhỏ. Phương pháp của họ đã không được áp dụng rộng rãi cho tới thập niên 1950, khi poliovirus được nuôi ở quy mô lớn phục vụ việc sản xuất vaccine.[22]
Một bước đột phá khác đến vào năm 1931, khi nhà bệnh học Hoa Kỳ Ernest William Goodpasture đã nuôi dưỡng virus cúm và vài loại virus khác trong trứng gà đã thụ tinh.[23] Năm 1949, John Franklin Enders, Thomas Weller, và Frederick Robbins cũng nuôi cấy virus bại liệt trong tế bào phôi người, virus đầu tiên được nuôi mà không sử dụng mô thể rắn của động vật hay trứng. Công trình này cho phép Jonas Salk tạo ra một vắc-xin bại liệt hiệu quả.[24]
Những hình ảnh đầu tiên của virus thu nhận được là nhờ sự phát minh ra kính hiển vi điện tử năm 1931 của hai kĩ sư người Đức Ernst Ruska và Max Knoll.[25] Năm 1935, Wendell Meredith Stanley - một nhà sinh hóa và virus học người Mỹ - đã nghiên cứu virus khảm thuốc lá và nhận thấy chúng được tạo thành phần lớn từ protein.[26] Một thời gian ngắn sau, virus này đã được phân tách thành các phần protein và ARN riêng biệt.[27] Virus khảm thuốc lá là dạng virus đầu tiên được tinh thể hóa và cấu trúc của nó do đó đã được làm sáng tỏ chi tiết. Những hình ảnh nhiễu xạ tia X đầu tiên của virus kết tinh đã được Bernal và Fankuchen thu được vào 1941. Dựa trên những tấm hình này, Rosalind Franklin đã khám phá ra cấu trúc ADN hoàn thiện của loại virus này vào năm 1955.[28] Cùng trong năm đó, Heinz Fraenkel-Conrat và Robley Williams chứng minh được là chiết xuất ARN và vỏ protein của virus khảm thuốc lá có thể tự lắp ráp lại để tạo thành những virus có chức năng, cho thấy cơ chế đơn giản này có thể là cách virus sinh ra trong tế bào vật chủ.[29]
Nửa sau của thế kỷ 20 đánh dấu một kỉ nguyên vàng cho sự khám phá virus với hầu hết trong số hơn 2.000 loài virus động vật, thực vật và vi khuẩn đã được phát hiện trong những năm này.[30] Năm 1957, virus arteri ở ngựa và virus gây bệnh tiêu chảy ở bò (một loại pestivirus) đã đươc phát hiện. Năm 1963, virus viêm gan siêu vi B cũng được Baruch Blumberg khám phá,[31] và năm 1965, Howard Temin đã mô tả loại retrovirus đầu tiên. Sau đó, enzym phiên mã ngược (Reverse transcriptase), loại enzym quan trọng mà retrovirus sử dụng để phiên mã ARN của chúng thành ADN, được miêu tả lần đầu vào năm 1970, một cách độc lập bởi Howard Martin Temin và David Baltimore.[32] Năm 1983, nhóm nghiên cứu của Luc Montagnier tại Viện Pasteur ở Pháp, đã lần đầu tiên phân lập một loại retrovirus được biết với tên gọi ngày nay là virus suy giảm miễn dịch ở người (HIV).[33]
Nguồn gốc
Virus được tìm thấy ở bất cứ nơi nào có sự sống và có thể đã tồn tại kể từ khi tế bào sống đầu tiên được tiến hóa thành.[34] Nguồn gốc của virus không rõ ràng bởi chúng không tạo hóa thạch, vì vậy các kĩ thuật phân tử đã được sử dụng để so sánh ARN hay ADN của virus và là một công cụ hiệu quả để nghiên cứu xem chúng phát sinh ra sao.[35] Nhìn chung, có ba giả thuyết chính để giải thích nguồn gốc của virus:[36][37]
Giả thuyết thoái lui (regressive hypothesis)
Giả thuyết này cho rằng virus có thể đã từng là những tế bào nhỏ mà ký sinh bên trong những tế bào lớn hơn. Trải qua thời gian, những gen không cần thiết cho sự ký sinh này mất đi. Những vi khuẩn như Rickettsia và Chlamydia cũng sống trong tế bào và giống như virus, chỉ có thể sinh sản khi ở bên trong tế bào vật chủ. Những vi khuẩn này đã hỗ trợ cho giả thuyết thoái lui, do sự phụ thuộc ký sinh của chúng có thể đã làm mất đi những gen cho phép chúng tồn tại bên ngoài tế bào. Giả thuyết này còn được gọi là "giả thuyết thoái hóa" (degeneracy hypothesis),[38][39] hoặc "giả thuyết suy giảm" (reduction hypothesis).[40] Giả thuyết nguồn gốc từ tế bào (Cellular origin hypothesis)
Theo giả thuyết này, một số virus có thể đã tiến hóa từ những mảnh ADN hay ARN mà "thoát ra" (escape) từ hệ gen của những sinh vật lớn hơn. ADN thoát ra có thể là từ những plasmid (những đoạn ADN trần mà có thể di chuyển giữa những tế bào) hoặc từ những transposon (những phân tử ADN mà nhân lên và di chuyển quanh những vị trí khác nhau bên trong bộ gen của tế bào).[41] Từng được gọi là những "gen nhảy", transposon là những ví dụ của các yếu tố di truyền di động và có thể là nguồn gốc của một số virus. Chúng được phát hiện ở ngô bởi Barbara McClintock vào năm 1950.[42] Đôi khi giả thuyết này còn được gọi là "giả thuyết lang thang" (vagrancy hypothesis),[38][43] hoặc "giả thuyết trốn thoát" (escape hypothesis).[40]
 [TẶNG BẠN] TRỌN BỘ Bí kíp học tốt 08 môn
[TẶNG BẠN] TRỌN BỘ Bí kíp học tốt 08 môn